Органоиды клетки
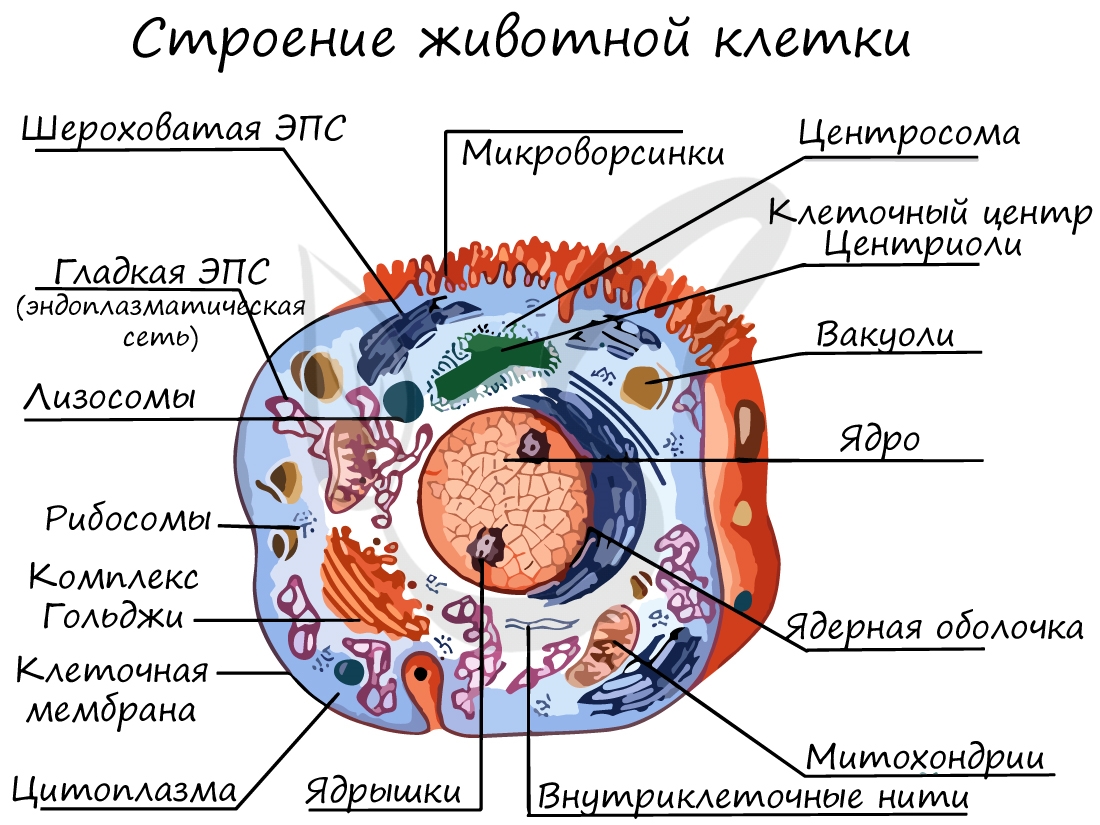
Клеточная мембрана (оболочка)
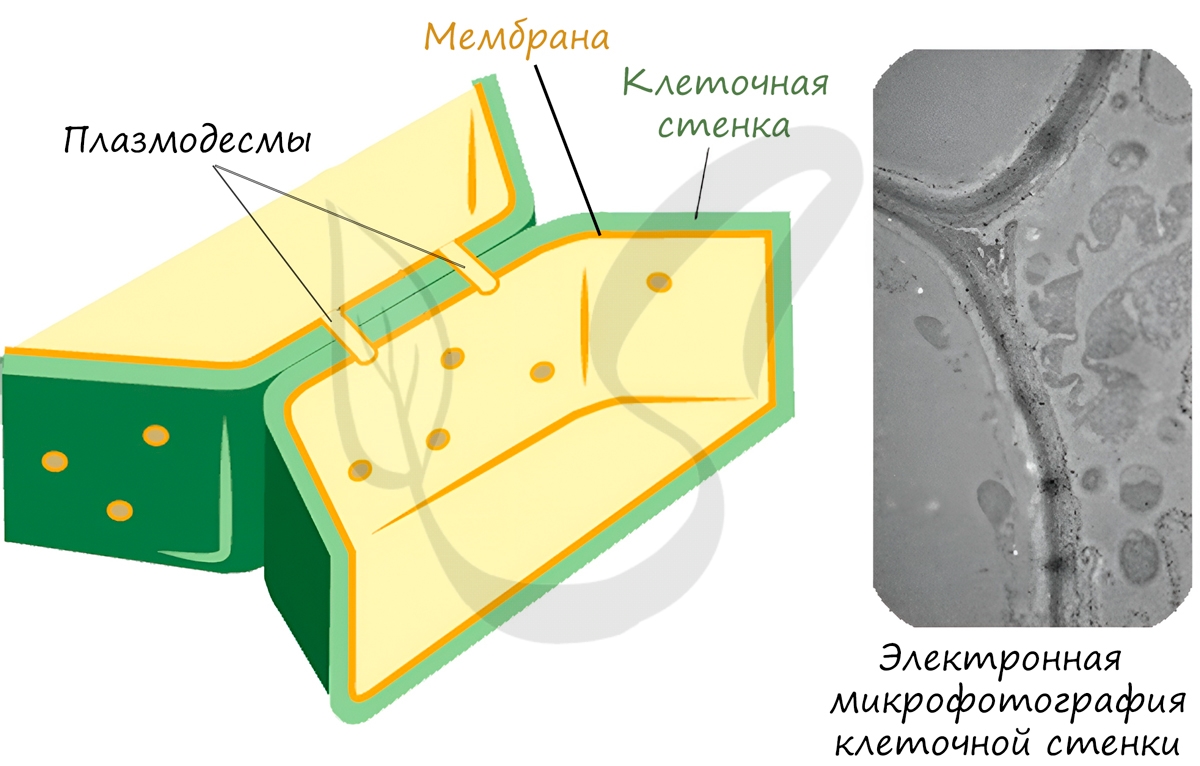
Запомните, что в отличие от клеточной стенки, которая есть только у растительных клеток и у клеток грибов (она придает им плотную, жесткую форму) клеточная мембрана есть у всех клеток без исключения! Этот важный момент объясню еще раз 🙂 У клеток животных имеется только клеточная мембрана, а у клеток растений и грибов есть и клеточная стенка, и клеточная мембрана.
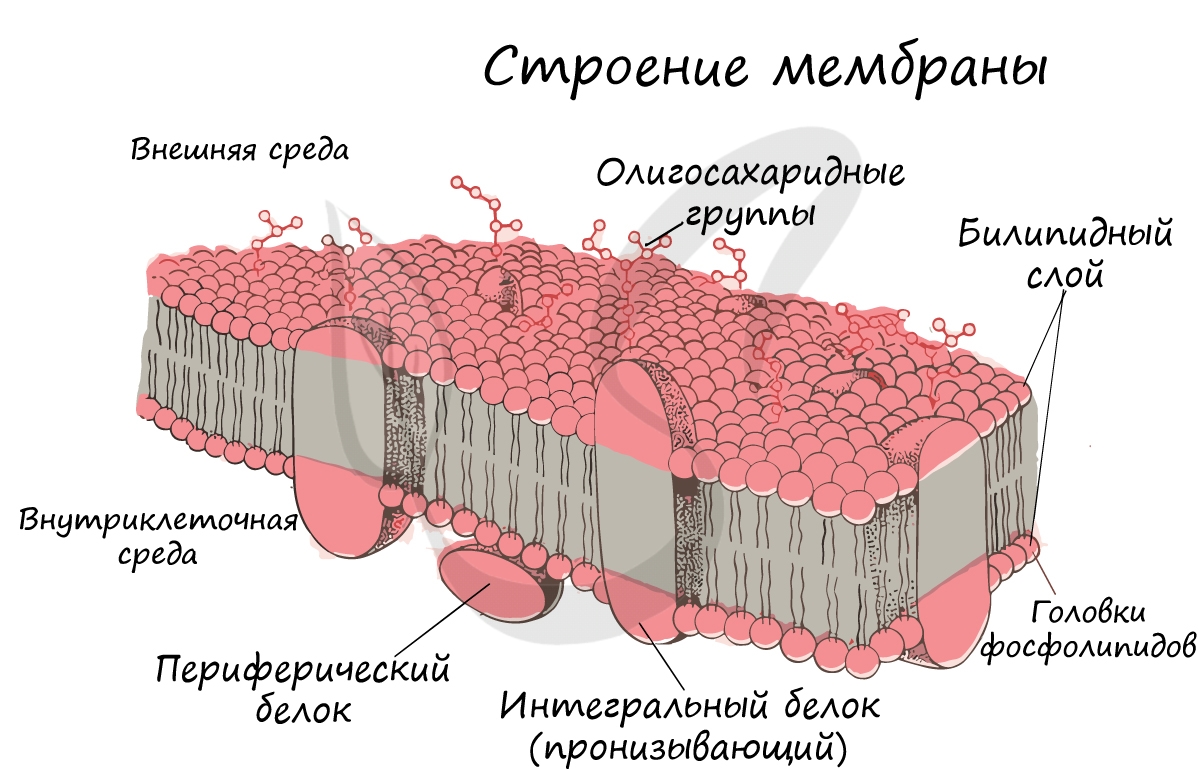
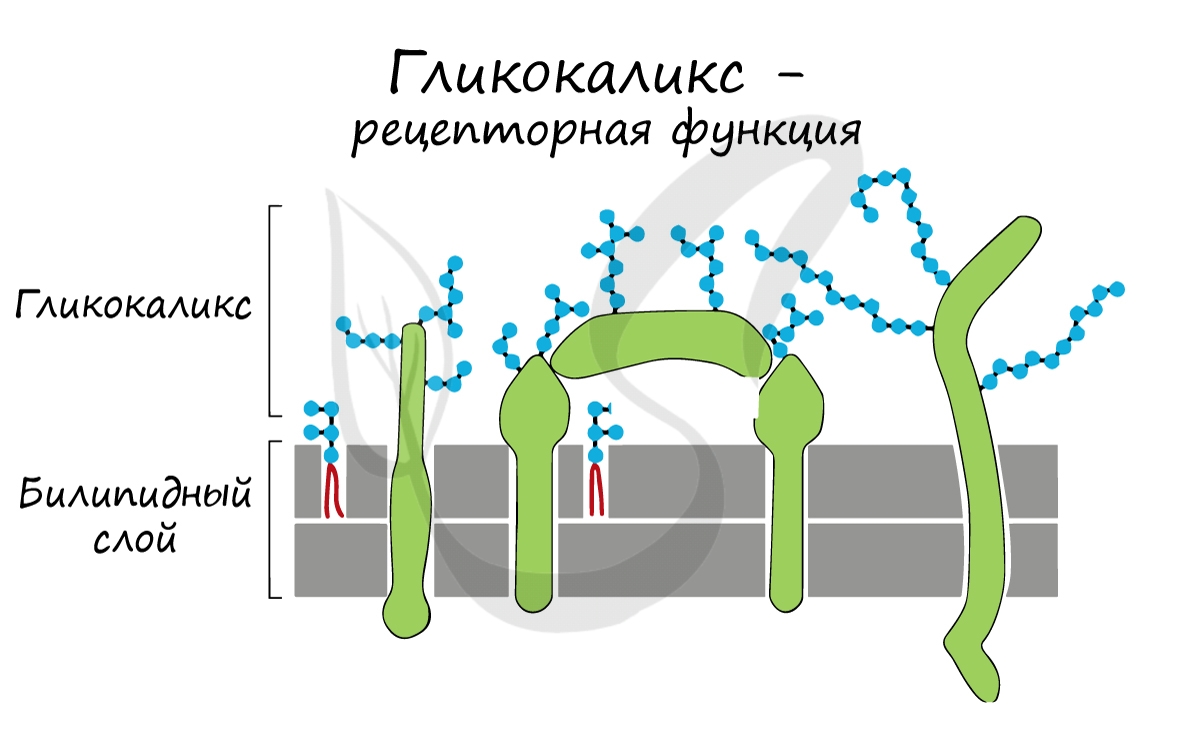
Интегральные (пронизывающие) белки образуют каналы, по которым молекулы различных веществ могут поступать в клетку или удаляться из нее. «Заякоренные» молекулы олигосахаридов на поверхности клетки образуют гликокаликс, который выполняет рецепторную функцию, участвует в избирательном транспорте веществ через мембрану.
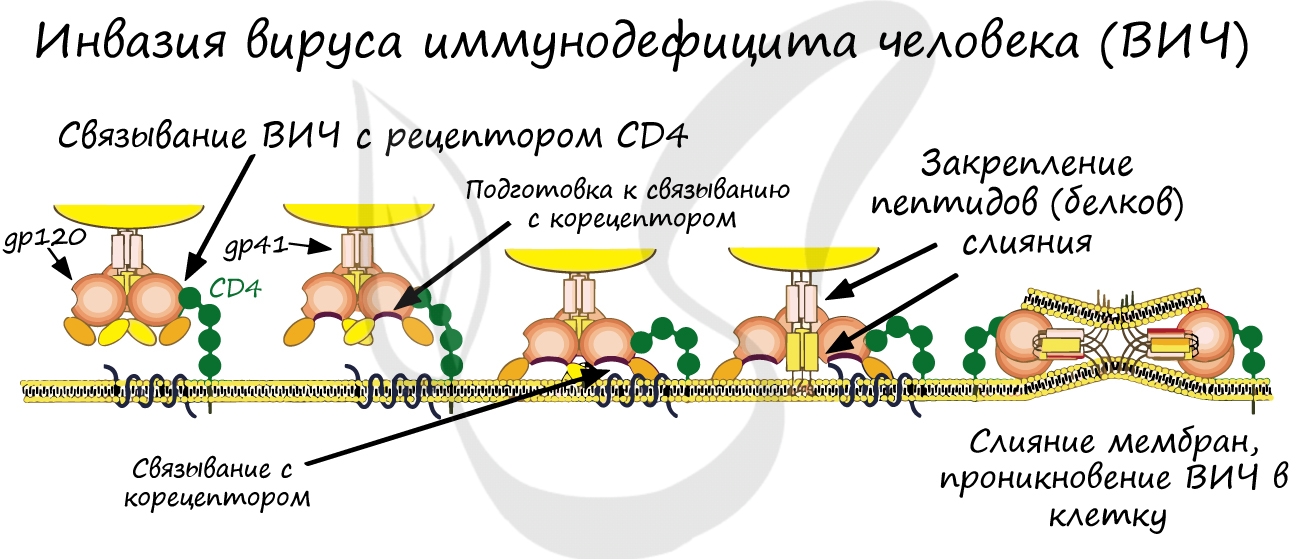
Вирусы и бактерии не являются исключением: они взаимодействуют только с теми клетками, на которых есть подходящие к ним рецепторы. Так, вирус гриппа поражает преимущественно клетки слизистой верхних дыхательных путей. Однако, если рецепторов нет, то вирус не может проникнуть в клетку, и организм приобретает невосприимчивость к инфекции. Вспомните врожденный иммунитет: именно по причине отсутствия рецепторов человек не восприимчив ко многим болезням животных.
Итак, вернемся к клеточной мембране. Ее можно сравнить со стенами помещения, в котором, вероятно, вы находитесь. Стены дома защищают его от ветра, дождя, снега и прочих факторов внешней среды. Рискну предположить, что в вашем доме есть окна и двери, которые по мере необходимости открываются и закрываются 🙂 Так и клеточная мембрана может сообщать внутреннюю среду клетки с внешней средой: через мембрану вещества поступают в клетку и удаляются из нее.
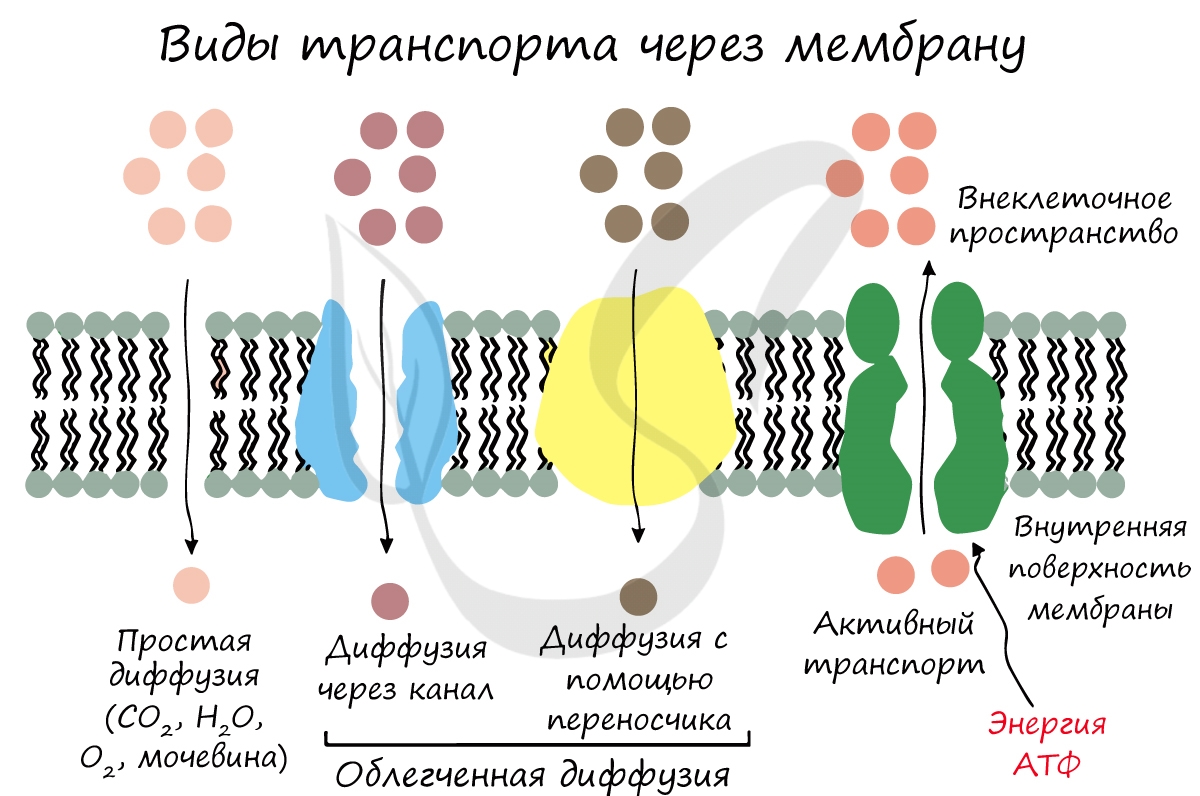
Внутрь клетки с помощью осмоса поступает вода. Путем простой диффузии в клетку попадают O2, H2O, CO2, мочевина. Облегченная диффузия характерна для транспорта глюкозы, аминокислот.
Активный транспорт чаще происходит против градиента концентрации, в ходе него используются белки-переносчики и энергия АТФ. Ярким примером является натрий-калиевый насос, который накачивает ионы калия внутрь клетки, а ионы натрия выводит наружу. Это происходит против градиента концентрации, поэтому без затрат энергии (АТФ) не обойтись.
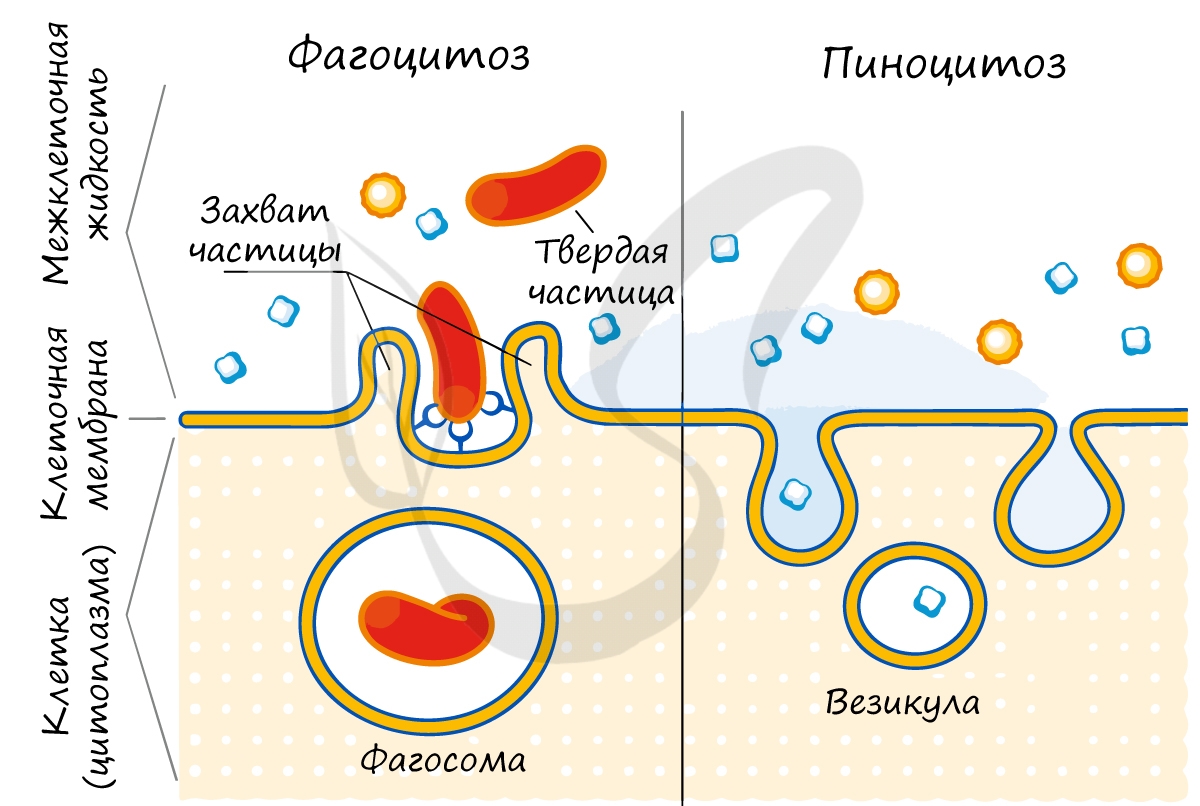
Фагоцитоз был открыт И.И. Мечниковым, который создал фагоцитарную теорию иммунитета. Это теория гласит, что в основе иммунной системы нашего организма лежит явление фагоцитоза: попавшие в организм бактерии уничтожаются фагоцитами (T-лимфоцитами), которые переваривают их.
В ходе эндоцитоза мембрана сильно прогибается внутрь клетки, ее края смыкаются, захватывая бактерию, пищевые частицы или жидкость внутрь клетки. Образуется везикула (пузырек), который движется к пищеварительной вакуоли или лизосоме, где происходит внутриклеточное пищеварение.
Клеточная стенка
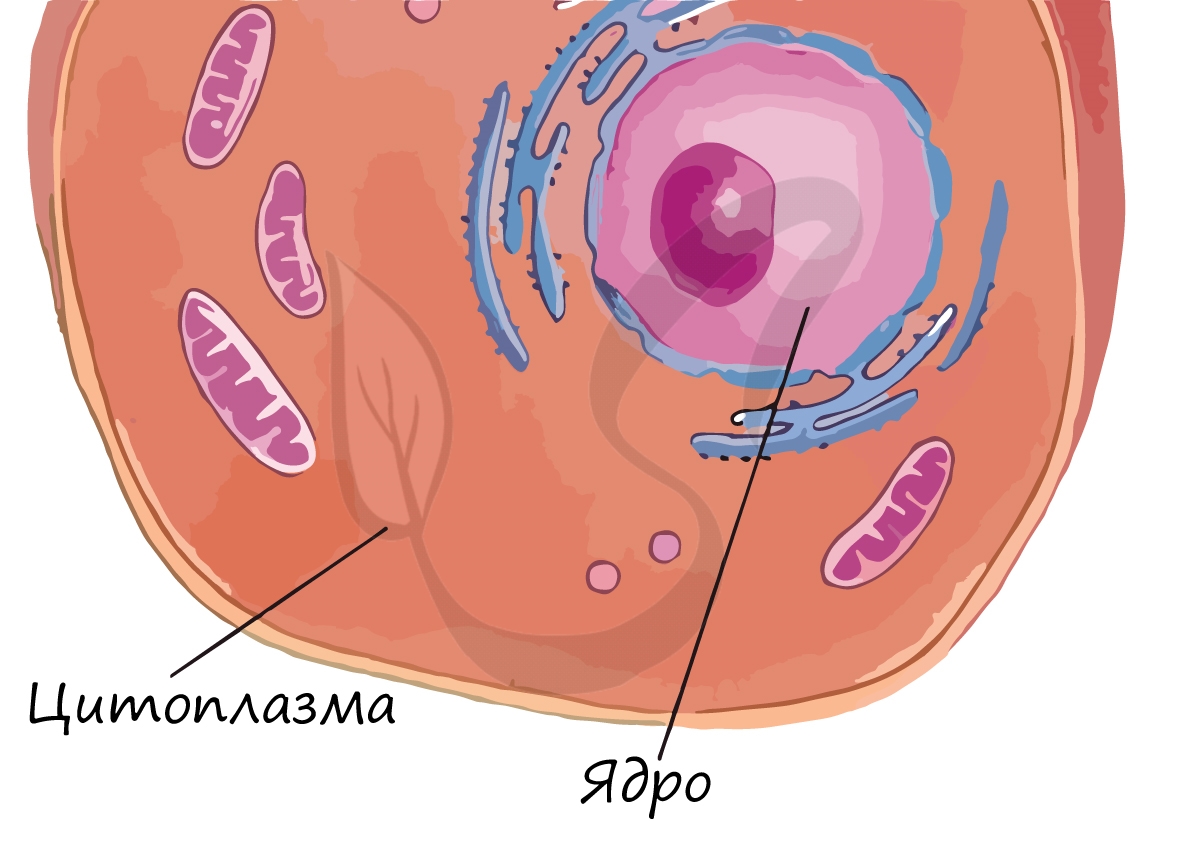
Цитоплазма
Постоянное движение цитоплазмы поддерживает связь между органоидами клетки и обеспечивает ее целостность.
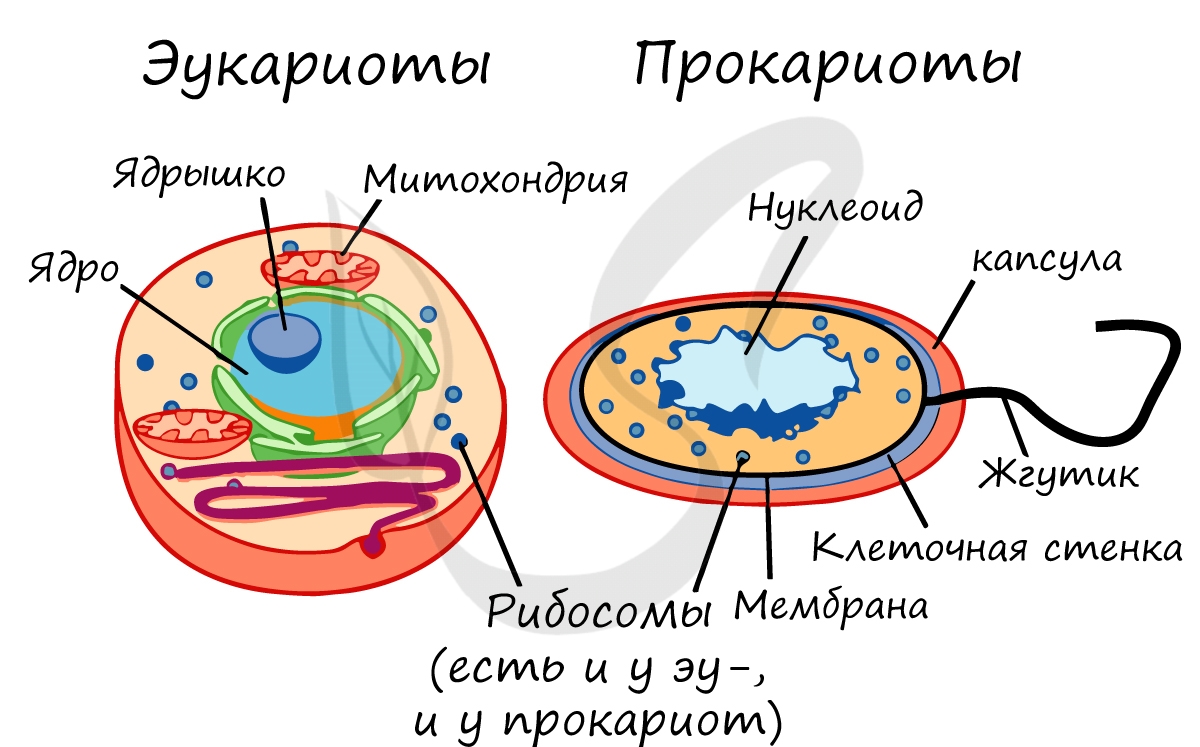
Прокариоты и эукариоты
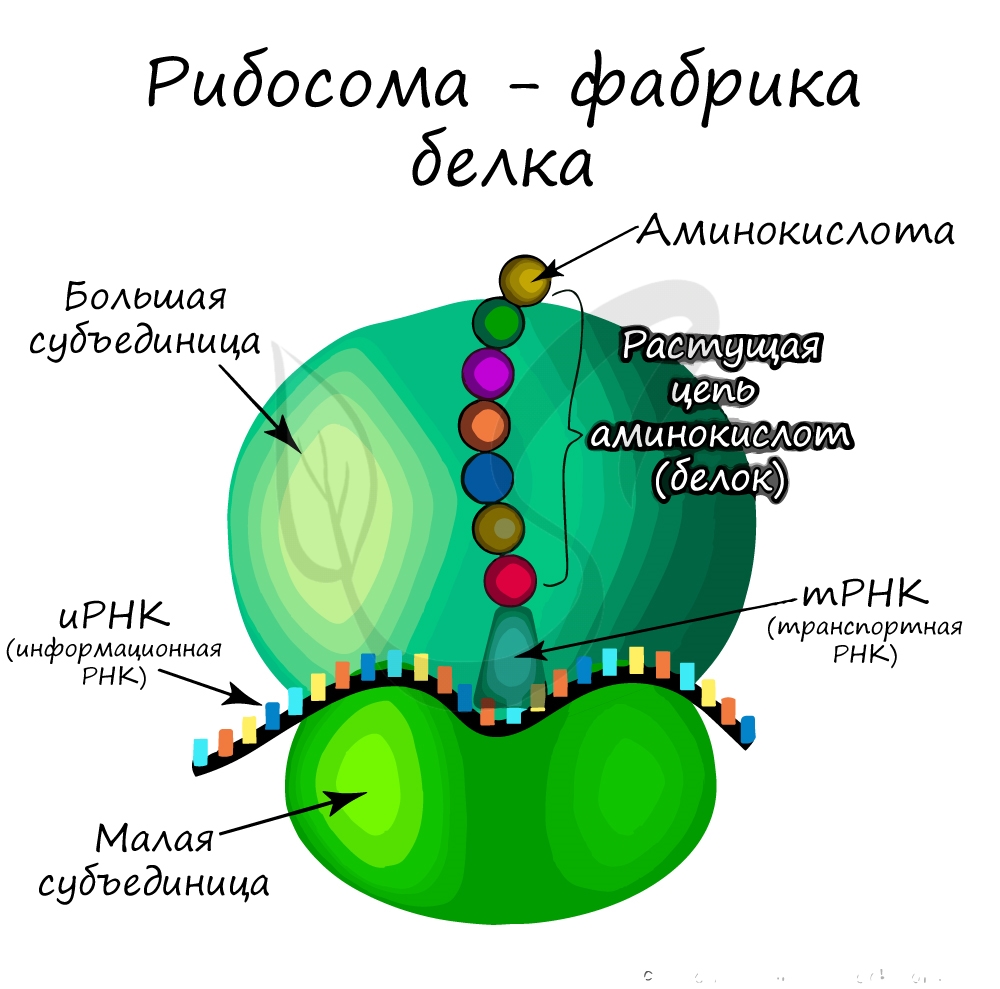
Немембранные органоиды
Очень мелкая органелла (около 20 нм), которая была открыта после появления электронного микроскопа. Состоит из двух субъединиц: большой и малой, в состав которых входят белки и рРНК (рибосомальная РНК), синтезируемая в ядрышке.
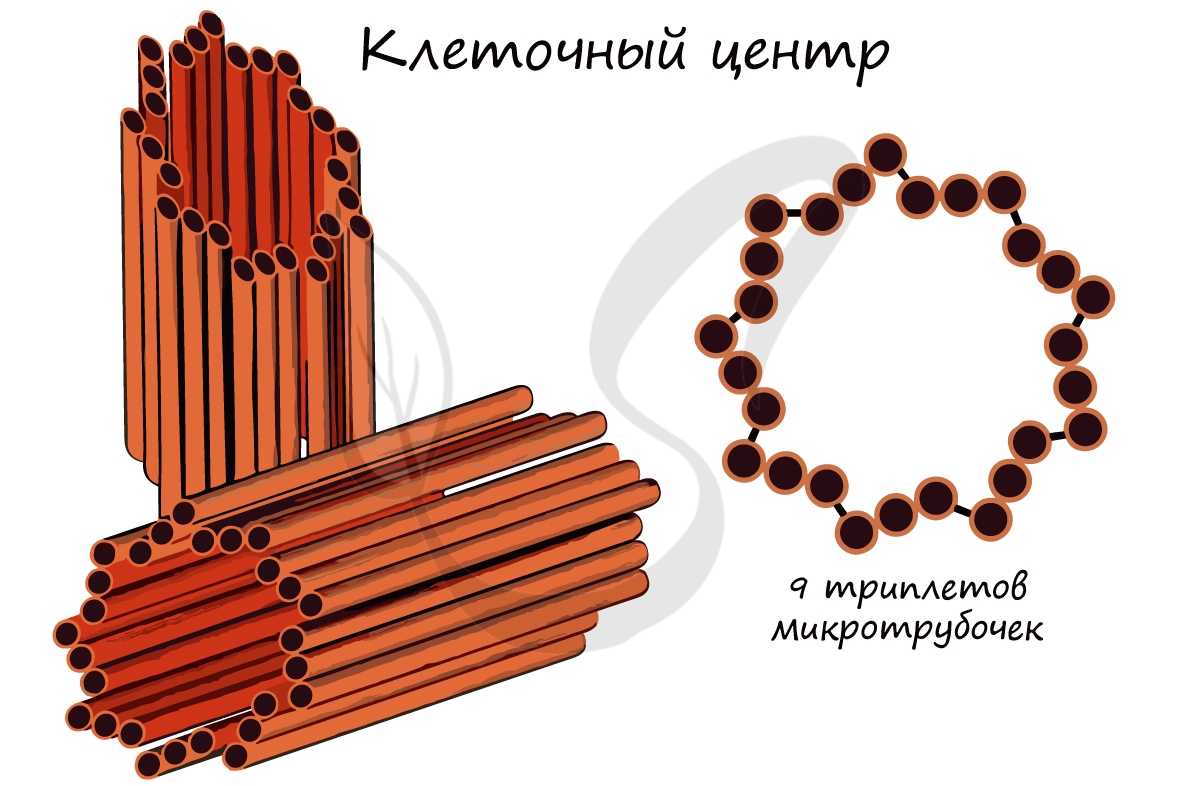
Это органоиды движения, которые выступают над поверхностью клетки и имеют в основе пучок микротрубочек. Реснички встречаются только в клетках животных, жгутики можно обнаружить у животных, растений и бактерий.
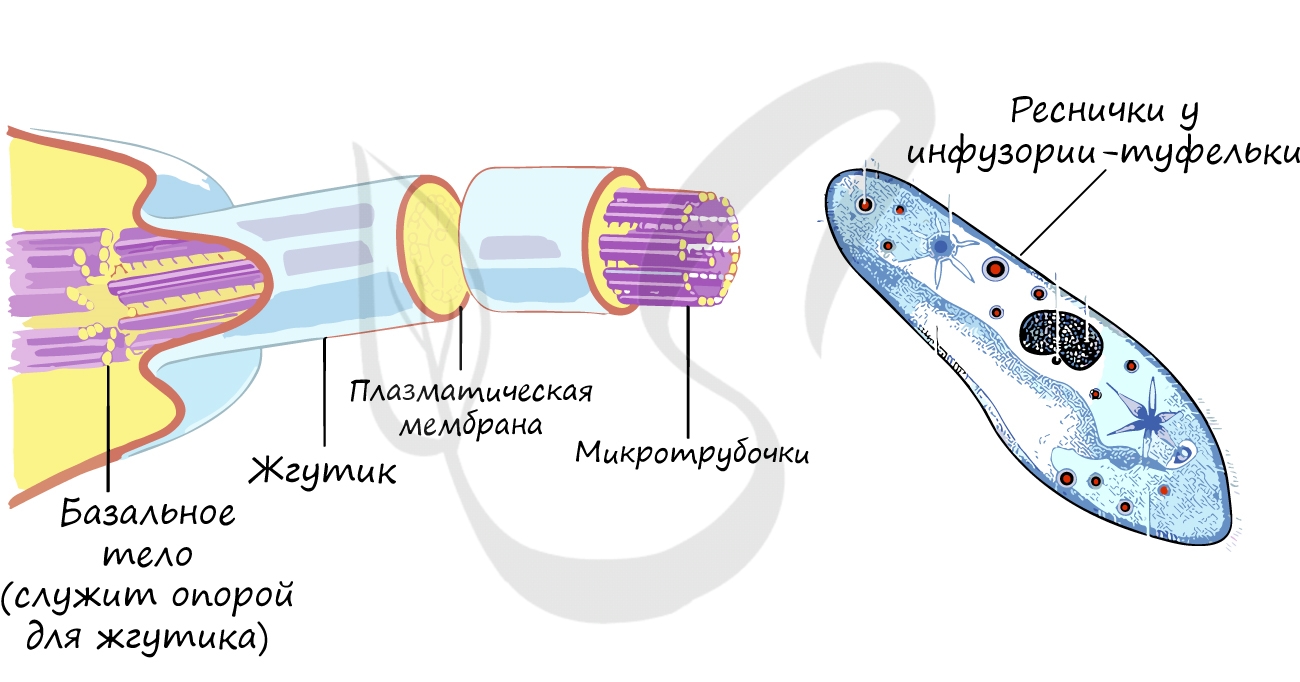
Одномембранные органоиды
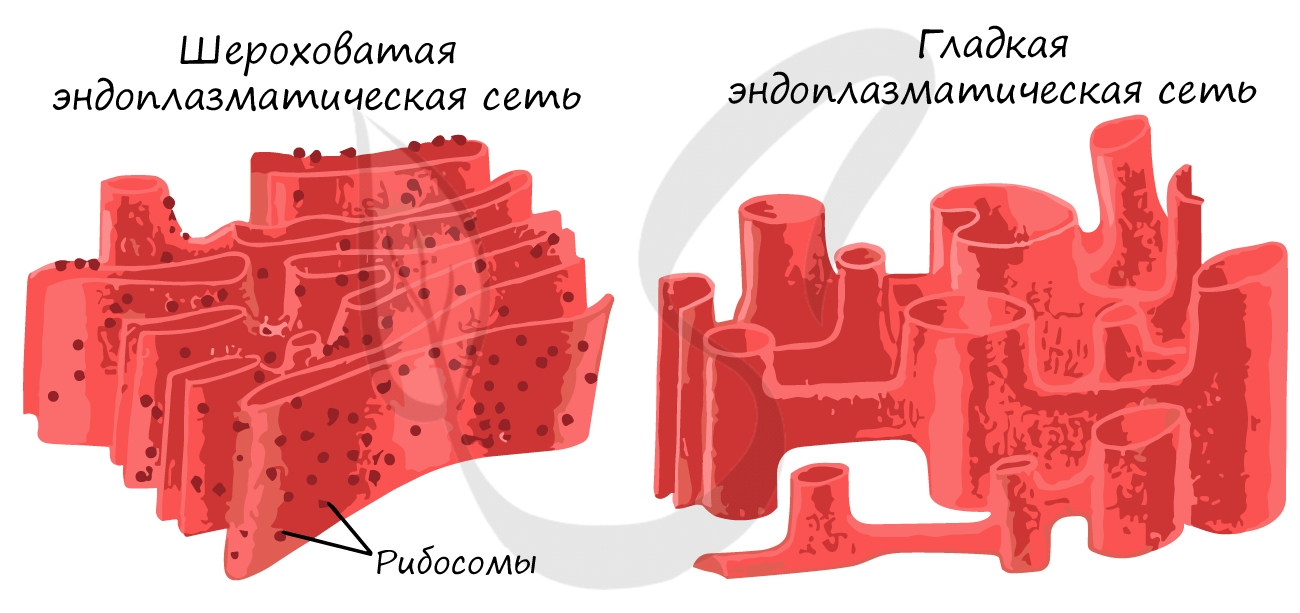
ЭПС представляет собой систему мембран, пронизывающих всю клетку и разделяющих ее на отдельные изолированные части (компартменты). Это крайне важно, так как в разных частях клетки идут реакции, которые могут помешать друг другу, что нарушит процессы жизнедеятельности.
Выделяют гладкую ЭПС и шероховатую ЭПС. Обе они выполняют функцию внутриклеточного транспорта веществ, однако между ними имеются различия. На мембранах гладкой ЭПС происходит синтез липидов, обезвреживаются вредные вещества. Шероховатая ЭПС синтезирует белок, так как имеет на мембранах многочисленные рибосомы (потому и называется шероховатой).
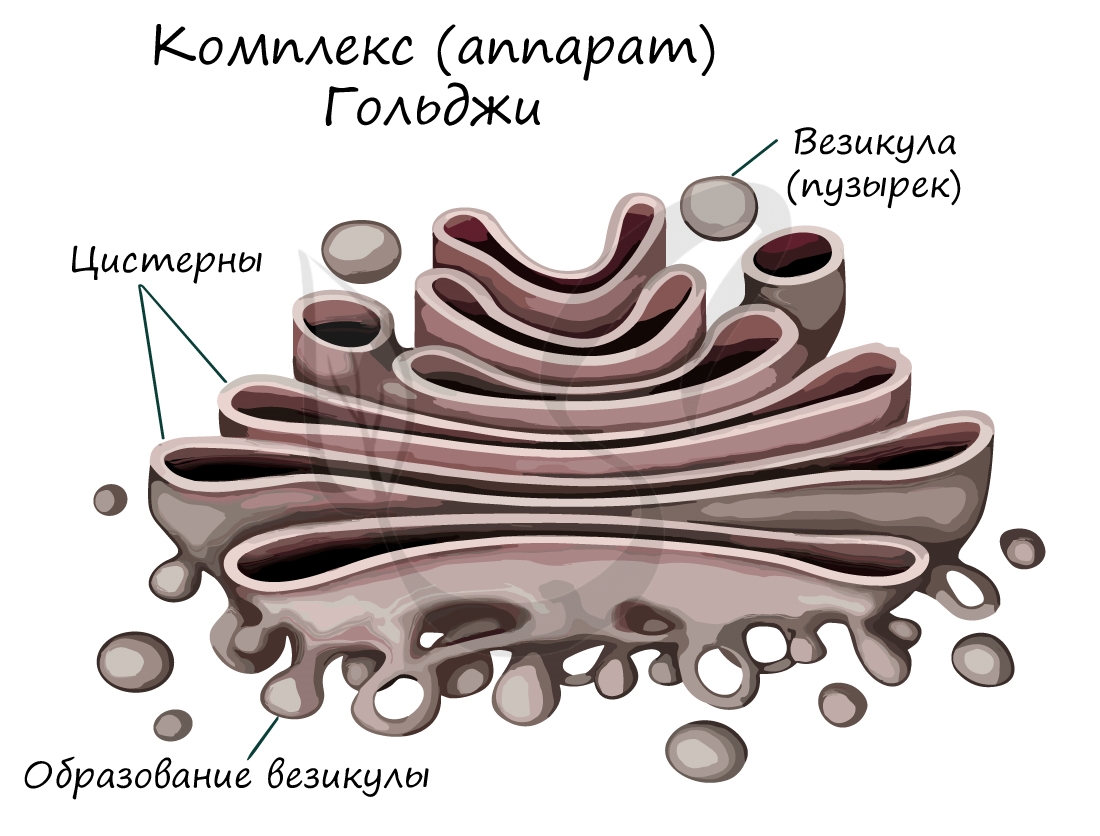
Модифицированные вещества упаковываются в пузырьки и могут перемещаться к мембране клетки, соединяясь с ней, они изливают свое содержимое во внешнюю среду. Можно догадаться, что комплекс Гольджи хорошо развит в клетках эндокринных желез, которые в большом количестве синтезируют и выделяют в кровь гормоны.
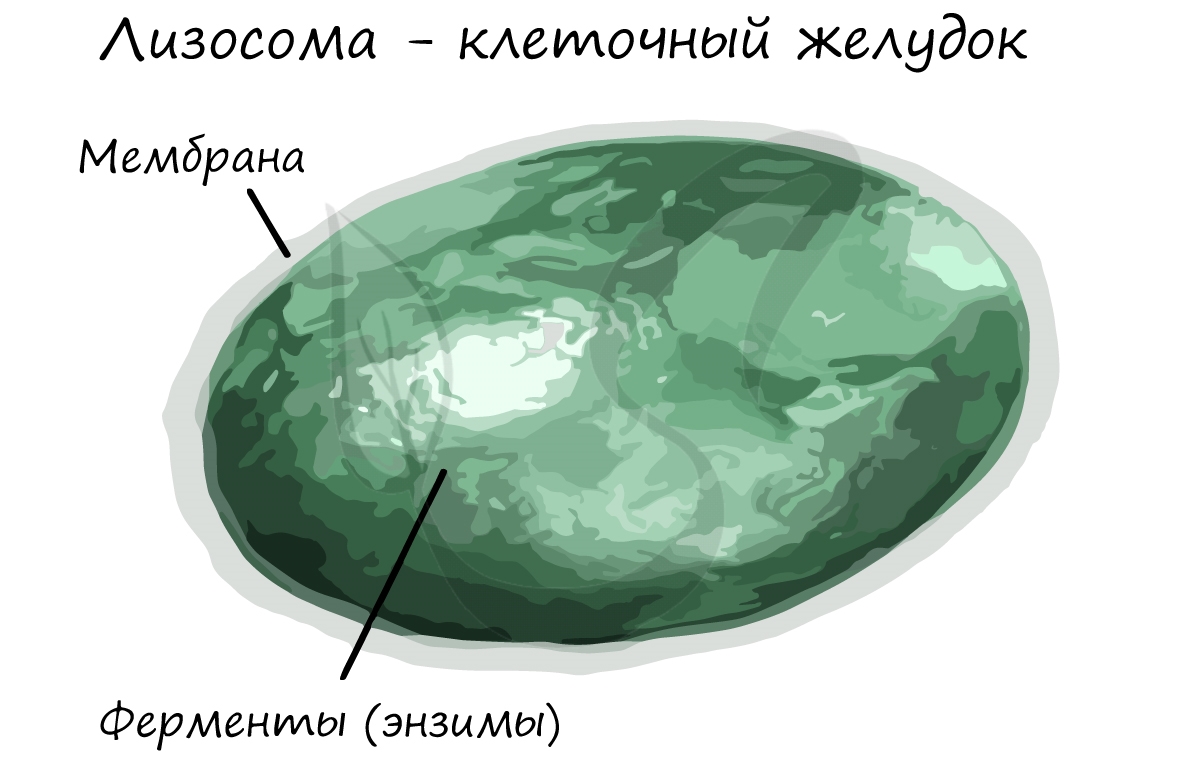
В комплексе Гольджи появляются первичные лизосомы, которые содержат ферменты в неактивном состоянии.

В ходе апоптоза ферменты лизосомы изливаются внутрь клетки, ее содержимое переваривается. Предполагают, что нарушение апоптоза в раковых клетках ведет к бесконтрольному росту опухоли.
Пероксисомы (микротельца) содержат окислительно-восстановительные ферменты, которые разлагают H2O2 (пероксид водорода) на воду и кислород. Если бы пероксид водорода оставался неразрушенными, это приводило бы к серьезным повреждениям клетки.
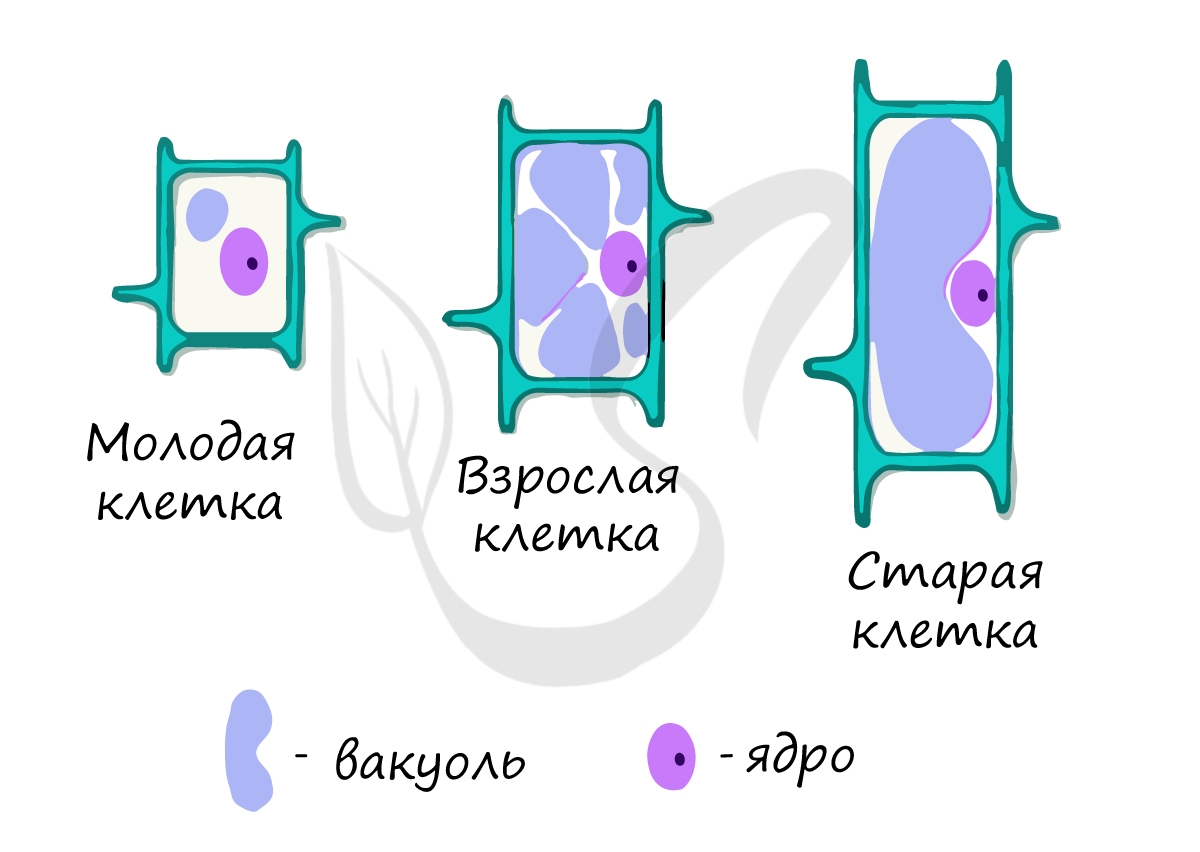
Трудно переоценить значение вакуолей в жизнедеятельности растительной клетки. Вакуоли создают осмотическое давление, придают клетке форму.
Примечательно, что по размеру вакуолей можно судить о возрасте клетки: молодые клетки имеют вакуоли небольшого размера, а в старых клетках вакуоли могут настолько увеличиваться, что оттесняют ядро и остальные органоиды на периферию.
Двумембранные органоиды
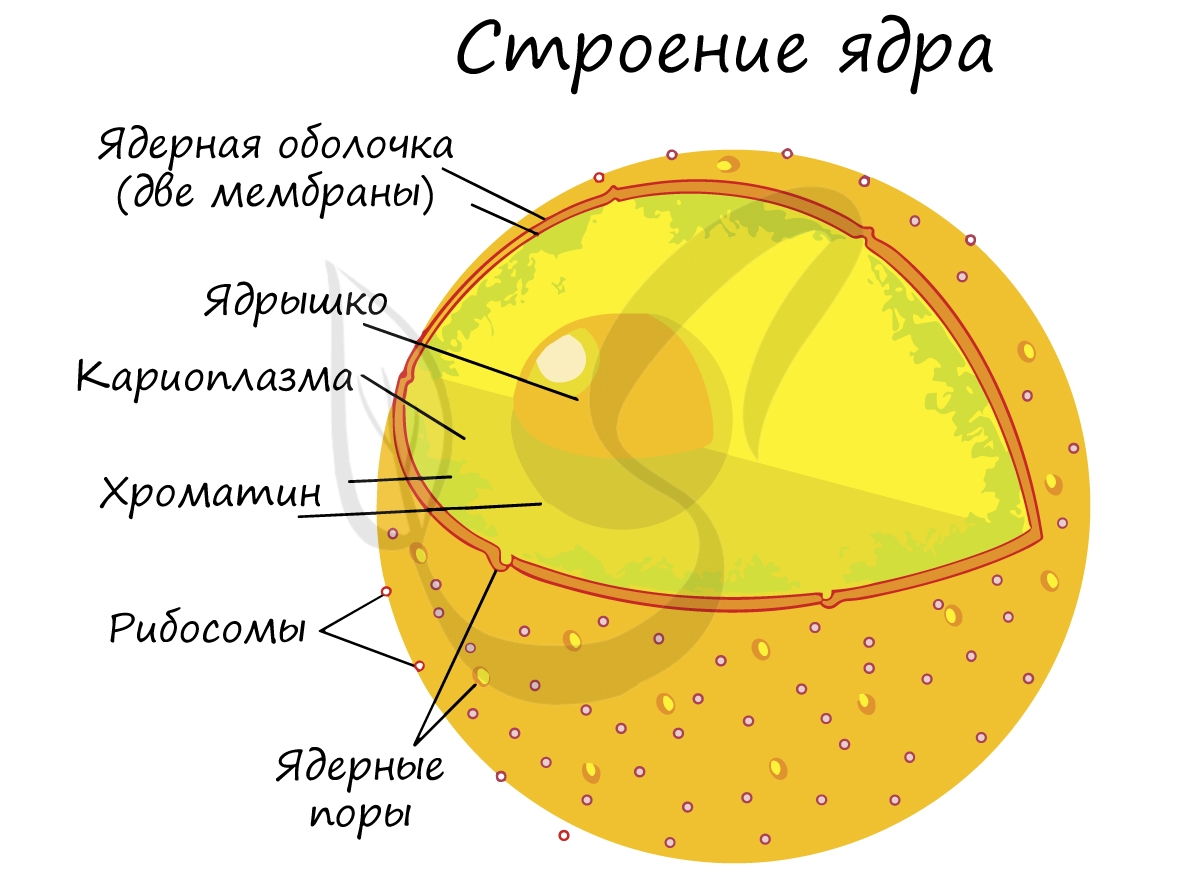
Оболочка ядра состоит из двух мембран и пронизана большим количеством ядерных пор, через которые происходит сообщение между кариоплазмой и цитоплазмой. Главными функциями ядра является хранение, защита и передача наследственного материала дочерним клеткам.
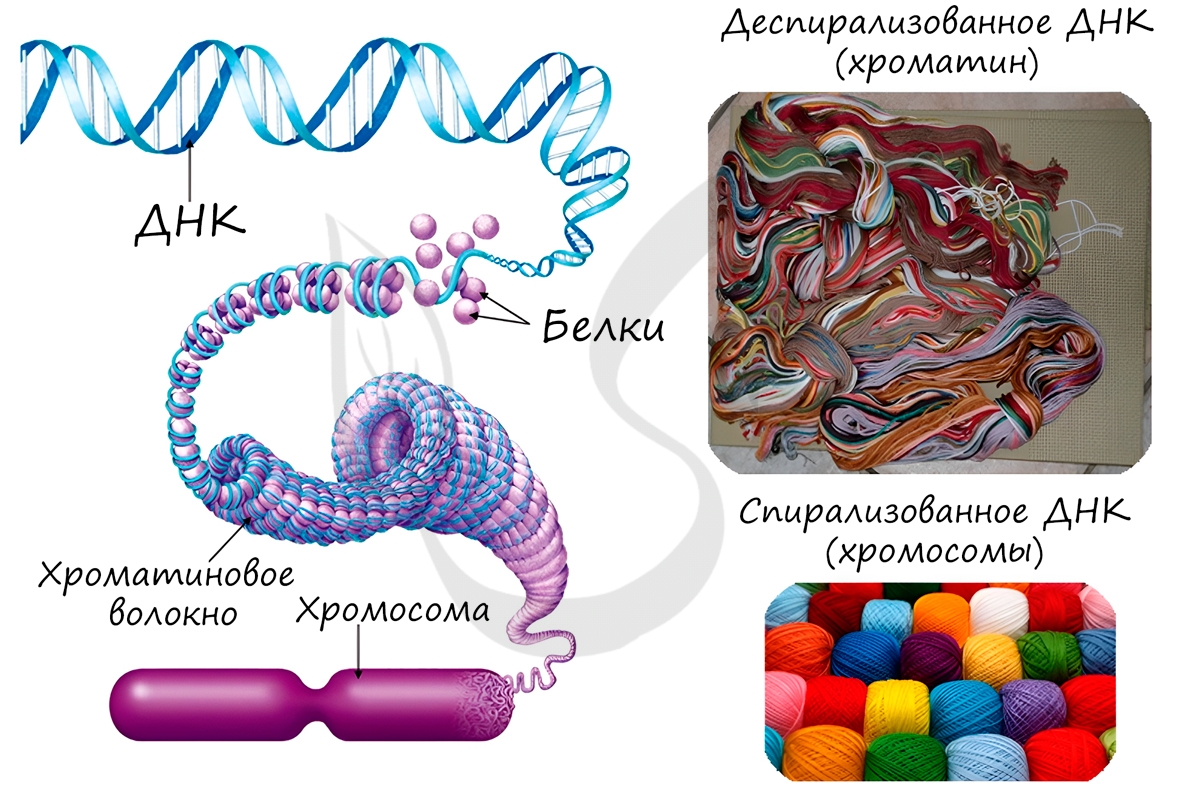
Замечу, что хромосомы видны только в момент деления клетки. Хромосомы представляют собой сильно спирализованные молекулы ДНК, связанные с белками.
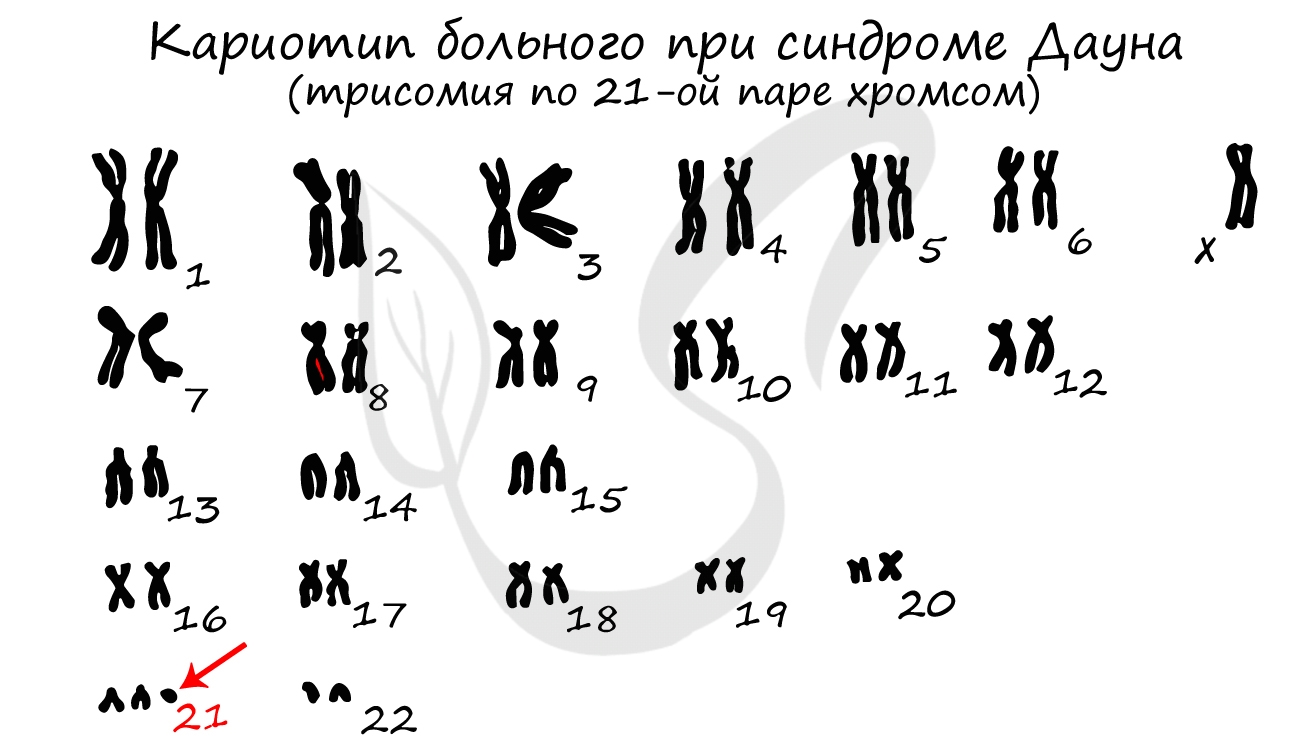
Хромосомы отличаются друг от друга по строению, форме, размерам. Совокупность всех признаков (форма, число, размер) хромосом называется кариотип. Кариотип может быть представлен по-разному: существует кариотип вида, особи, клетки.
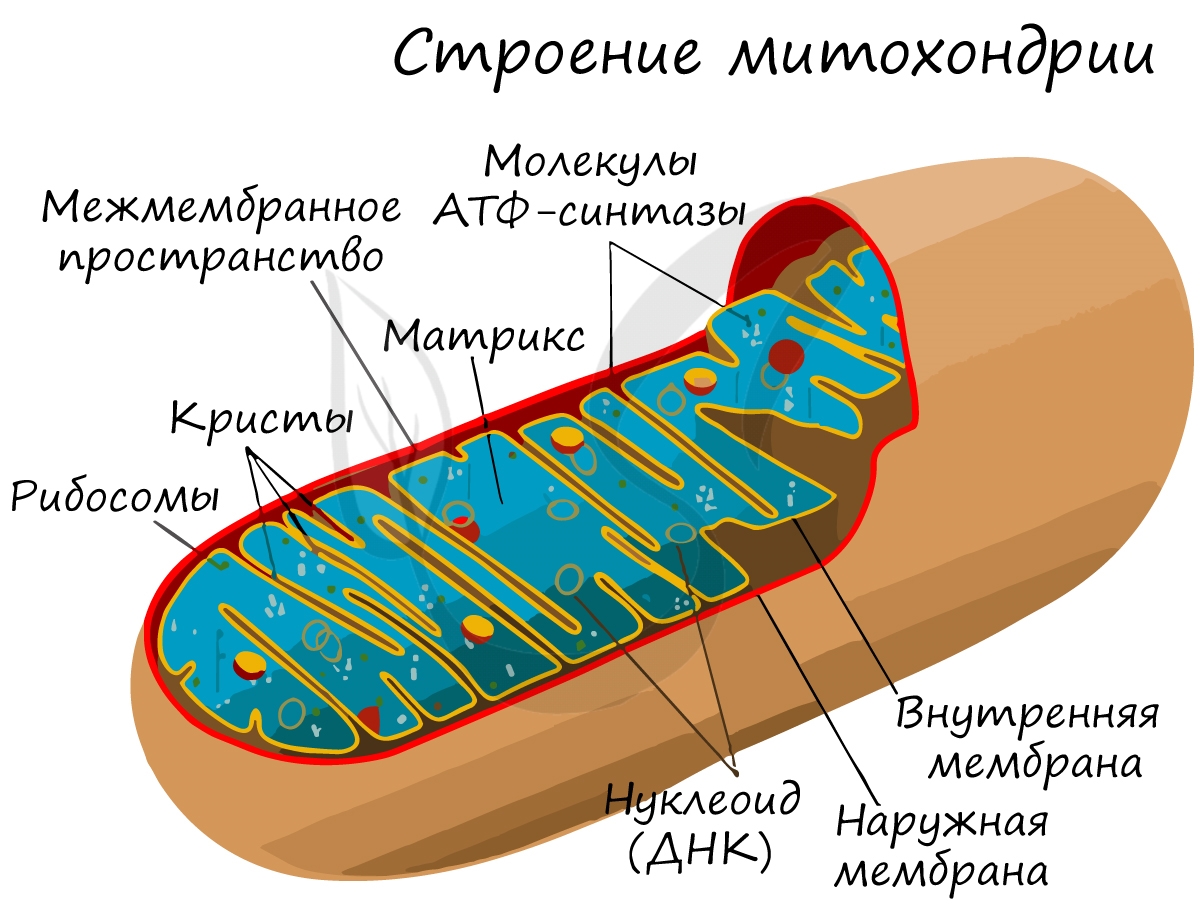
В связи с этим, митохондрия считается полуавтономным органоидом. Вероятнее всего, изначально митохондрии были самостоятельными организмами, однако со временем вступили в симбиоз с эукариотами и стали частью клетки.
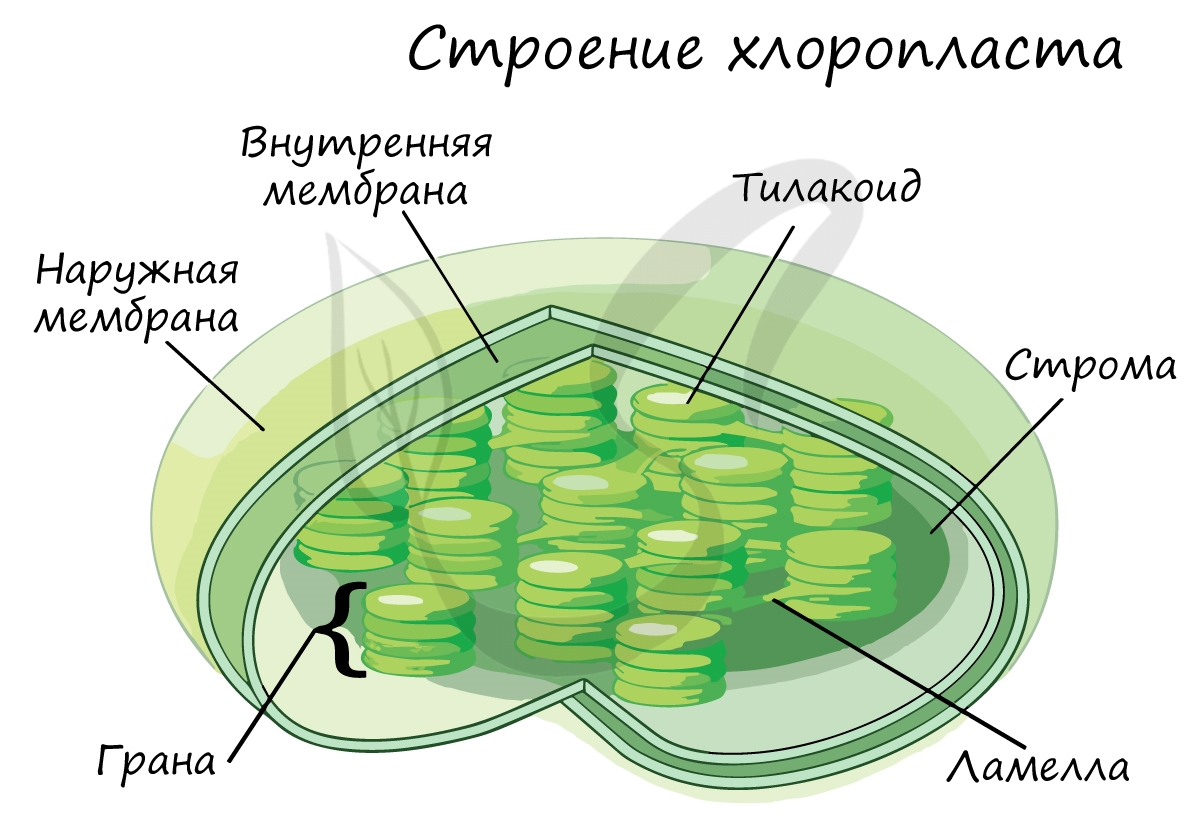
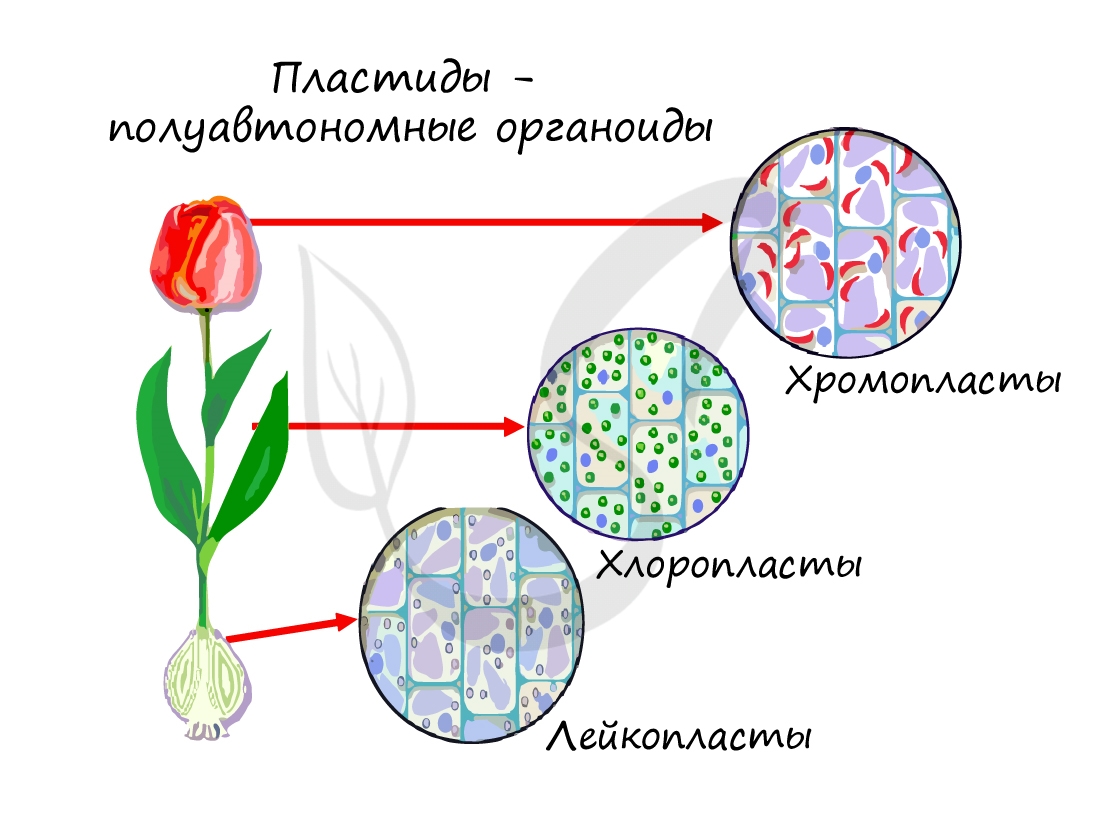
Так же, как и митохондрии, пластиды относятся к полуавтономным органоидам: в них имеется кольцевидная ДНК (находится в нуклеоиде), рибосомы.
Пластиды, которые содержат пигменты каратиноиды в различных сочетаниях. Сочетание пигментов обуславливает красную, оранжевую или желтую окраску. Находятся в плодах, листьях, лепестках цветков.
Хромопласты могут развиваться из хлоропластов: во время созревания плодов хлоропласты теряют хлорофилл и крахмал, в них активируется биосинтез каротиноидов.
Не содержат пигментов, образуются в запасающих частях растения (клубни, корневища). В лейкопластах накапливается крахмал, липиды (жиры), пептиды (белки). На свету лейкопласты могут превращаться в хлоропласты и запускать процесс фотосинтеза.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Транскрипция и трансляция

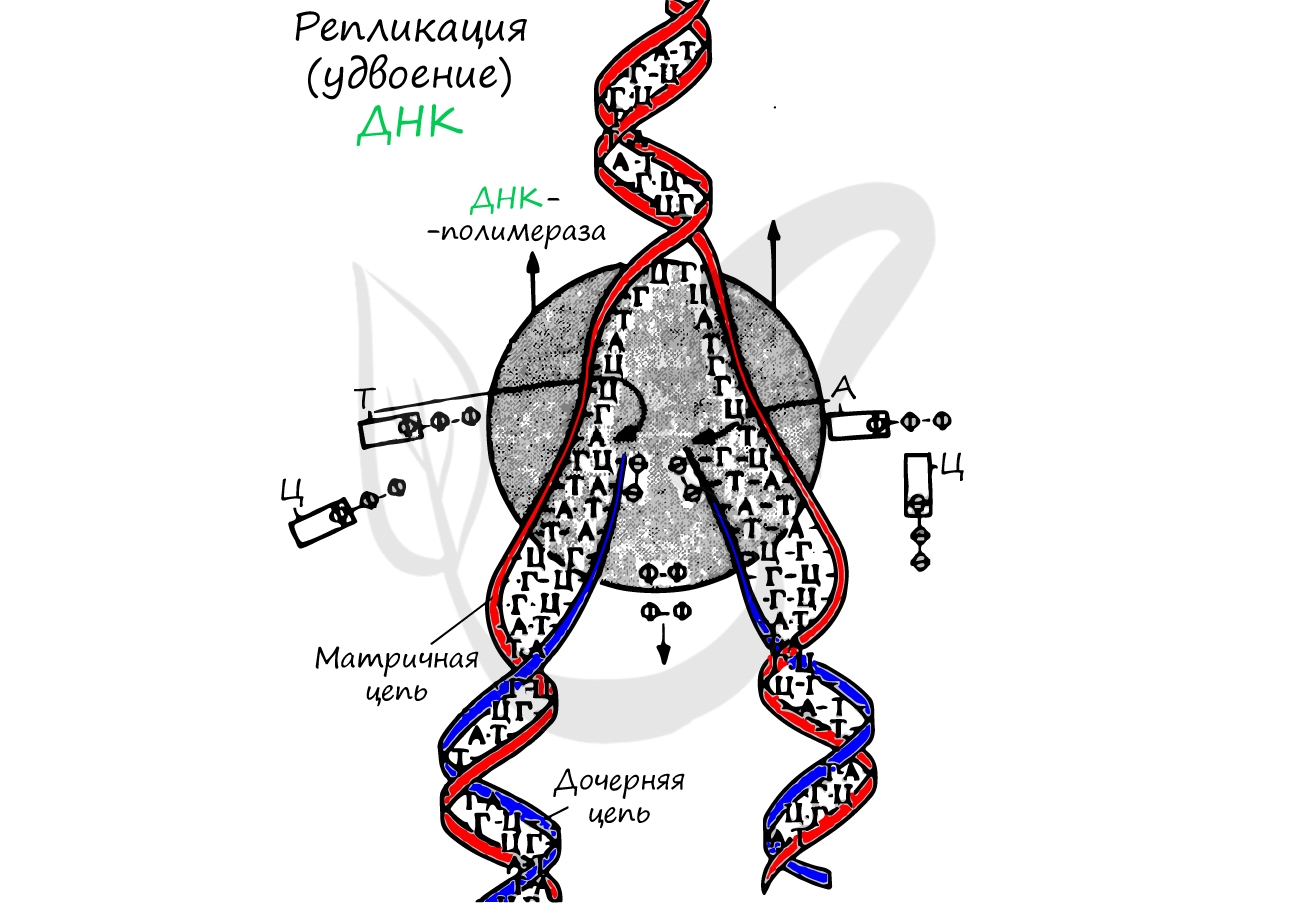
Удвоение ДНК происходит в синтетическом периоде интерфазы. При этом общее число хромосом не меняется, однако каждая из них содержит к началу деления две молекулы ДНК: это необходимо для равномерного распределения генетического материала между дочерними клетками.
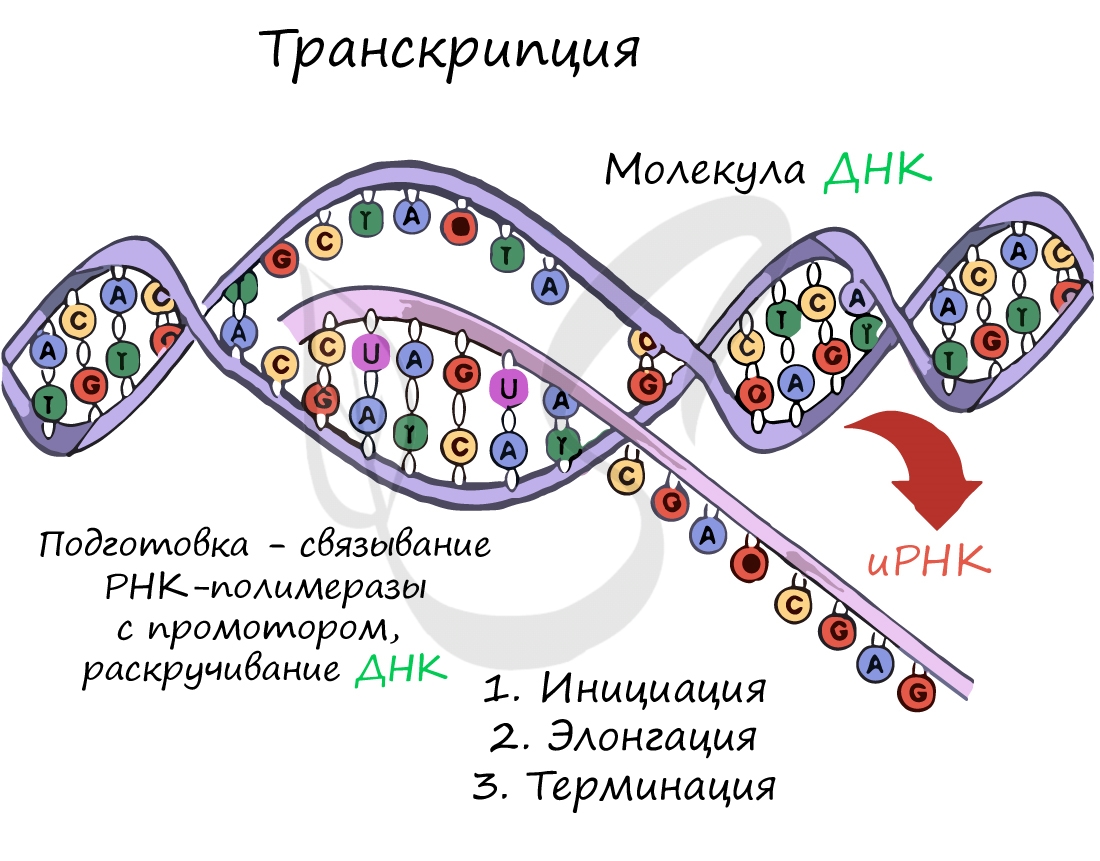
Транскрпиция (лат. transcriptio — переписывание)
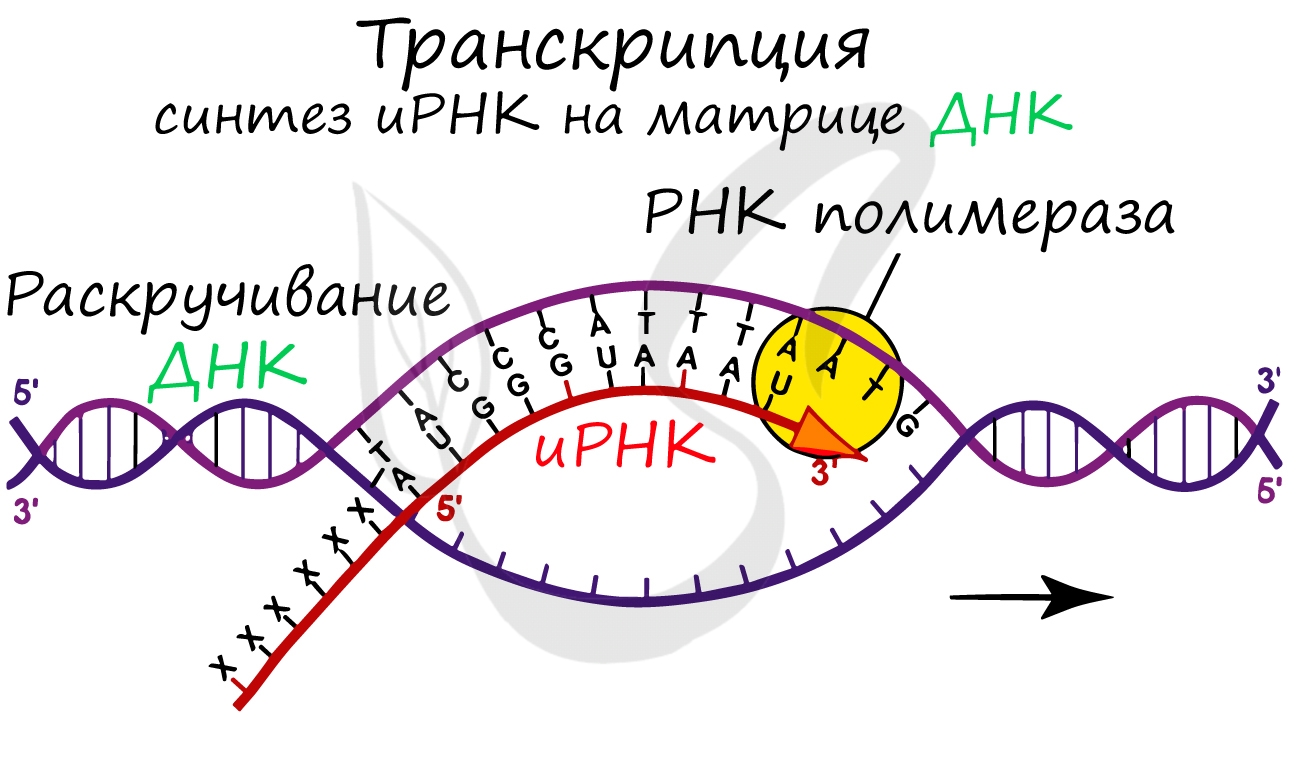
Образуется несколько начальных кодонов иРНК.
Нити ДНК последовательно расплетаются, освобождая место для передвигающейся РНК-полимеразы. Молекула иРНК быстро растет.
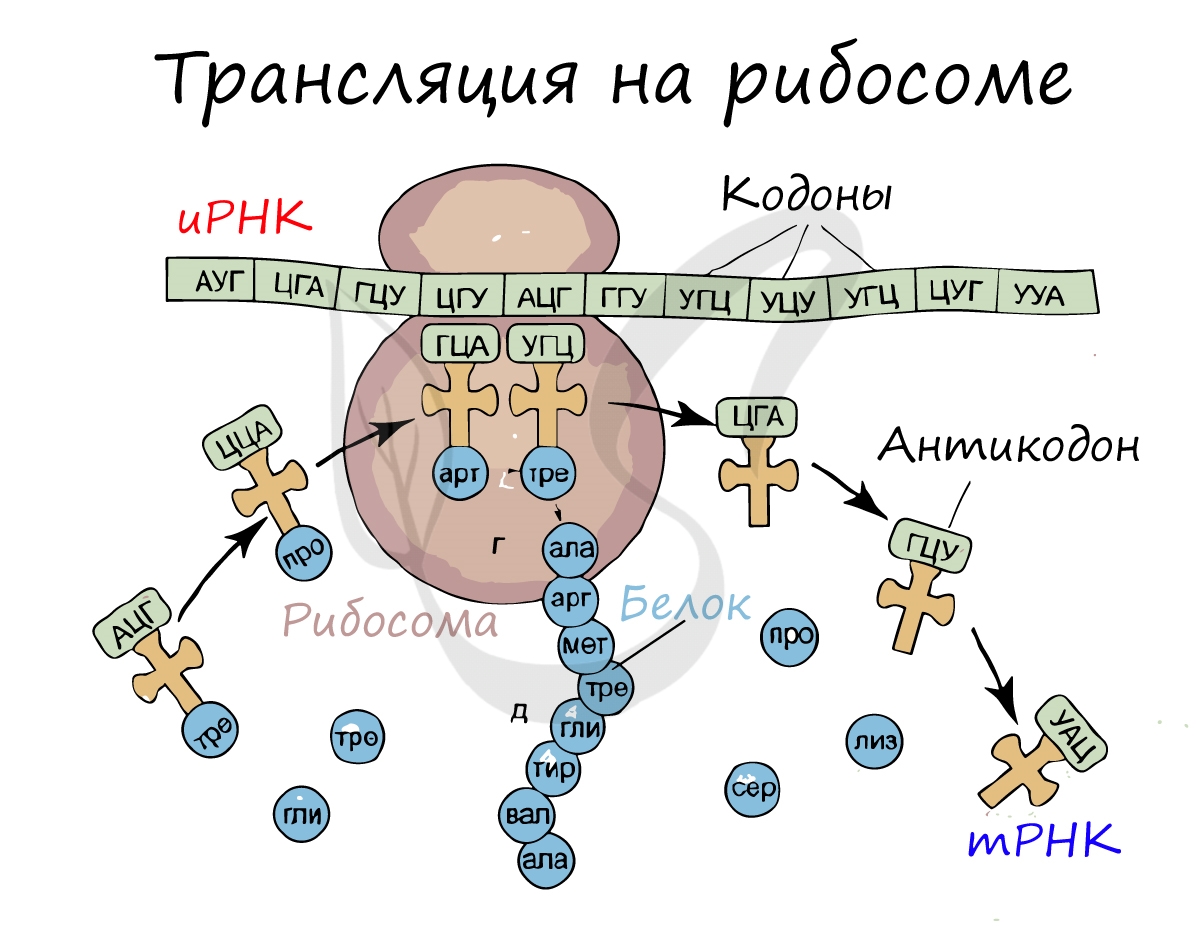
Трансляция (от лат. translatio — перенос, перемещение)
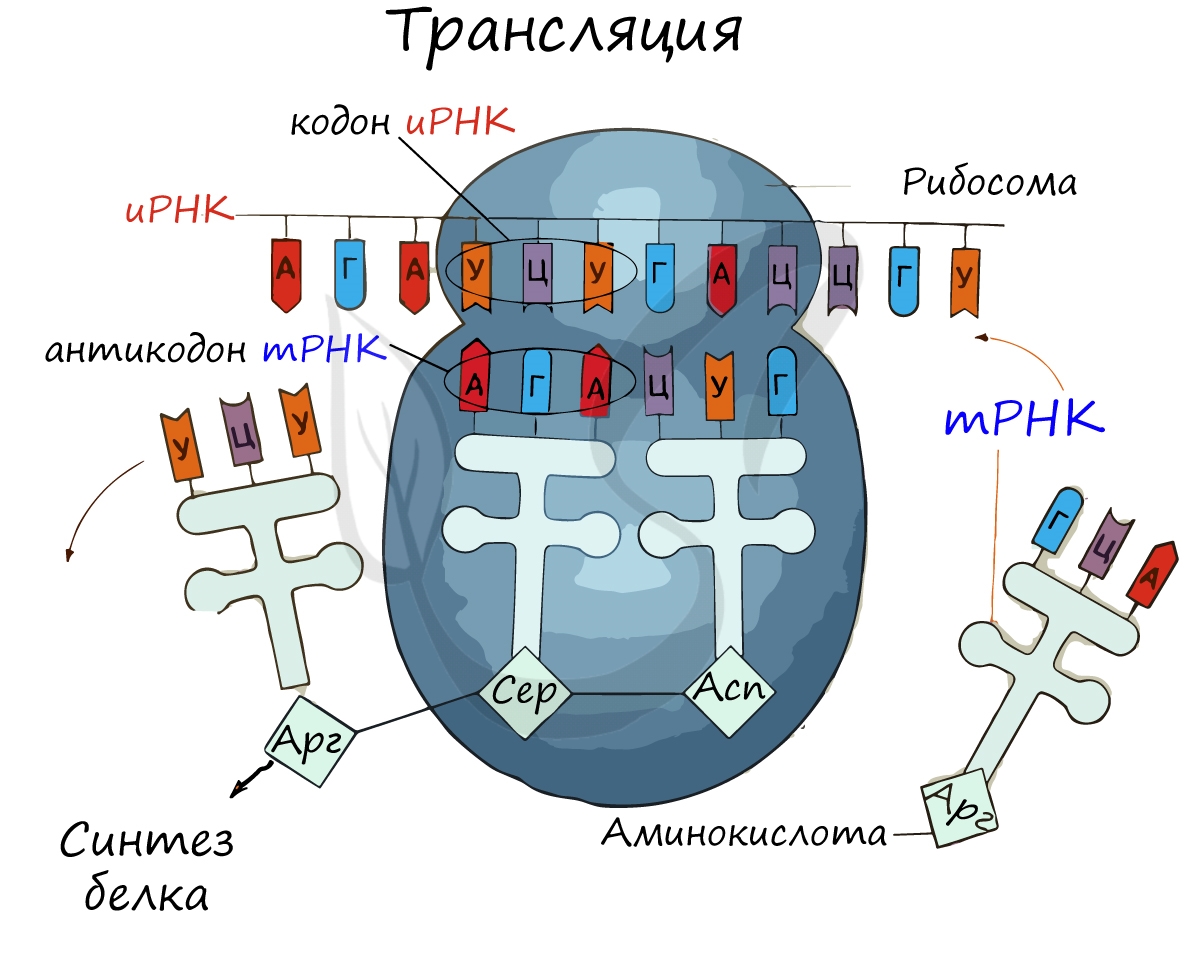
Рибосома делает шаг, и иРНК продвигается на один кодон: такое в фазу элонгации происходит десятки тысяч раз. Молекулы тРНК приносят новые аминокислоты, соответствующие кодонам иРНК. Аминокислоты соединяются друг с другом: между ними образуются пептидные связи, молекула белка растет.
Примеры решения задачи №1
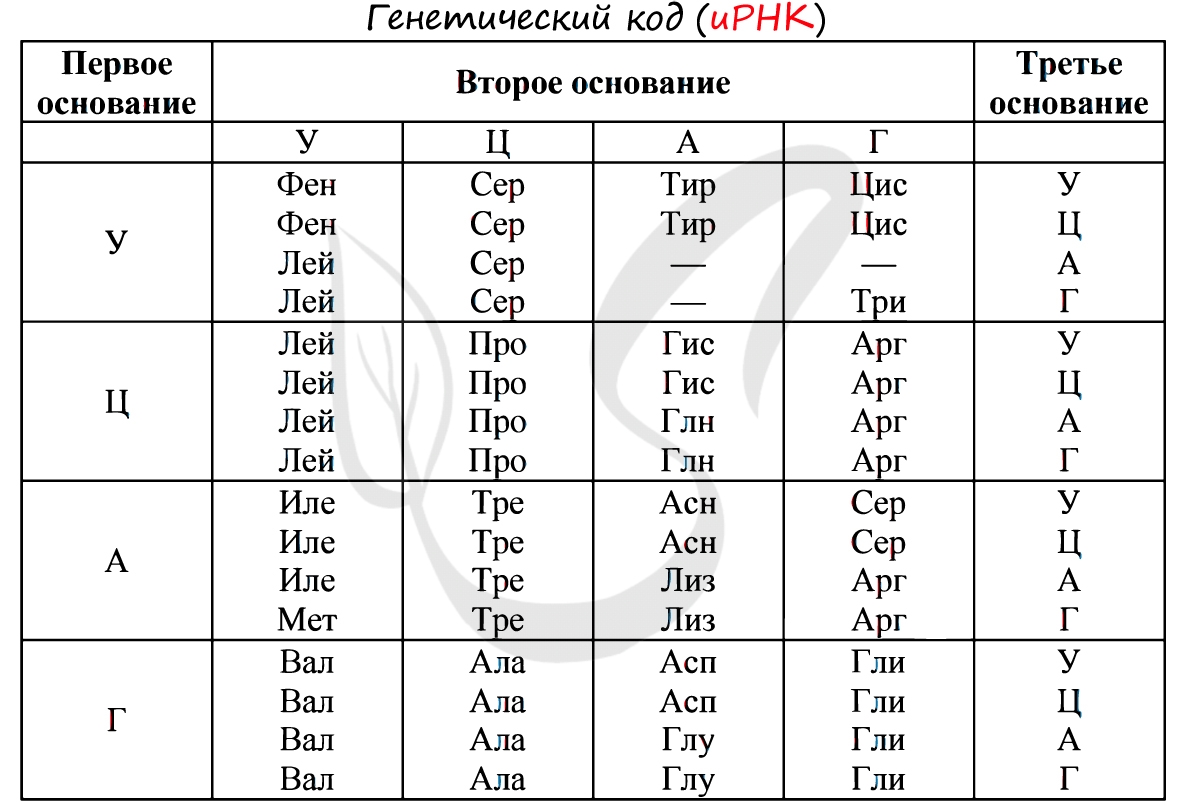
Без практики теория мертва, так что скорее решим задачи! В первых двух задачах будем пользоваться таблицей генетического кода (по иРНК), приведенной вверху.
«Фрагмент цепи ДНК имеет следующую последовательность нуклеотидов: ЦГА-ТГГ-ТЦЦ-ГАЦ. Определите последовательность нуклеотидов во второй цепочке ДНК, последовательность нуклеотидов на иРНК, антикодоны соответствующих тРНК и аминокислотную последовательность соответствующего фрагмента молекулы белка, используя таблицу генетического кода»

По принципу комплементарности мы нашли вторую цепочку ДНК: ГЦТ-АЦЦ-АГГ-ЦТГ. Мы использовали следующие правила при нахождении второй нити ДНК: А-Т, Т-А, Г-Ц, Ц-Г.
Вернемся к первой цепочке, и именно от нее пойдем к иРНК: ГЦУ-АЦЦ-АГГ-ЦУГ. Мы использовали следующие правила при переводе ДНК в иРНК: А-У, Т-А, Г-Ц, Ц-Г.
Зная последовательность нуклеотидов иРНК, легко найдем тРНК: ЦГА, УГГ, УЦЦ, ГАЦ. Мы использовали следующие правила перевода иРНК в тРНК: А-У, У-А, Г-Ц, Ц-Г. Обратите внимание, что антикодоны тРНК мы разделяем запятыми, в отличие кодонов иРНК. Это связано с тем, что тРНК представляют собой отдельные молекулы (в виде клеверного листа), а не линейную структуру (как ДНК, иРНК).
Пример решения задачи №2
«Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК, имеет следующую последовательность нуклеотидов: ТАГ-ЦАА-АЦГ-ГЦТ-АЦЦ. Установите нуклеотидную последовательность участка тРНК, который синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта тРНК в процессе биосинтеза белка, если третий триплет соответствует антикодону тРНК»

Пример решения задачи №3
Длина фрагмента молекулы ДНК составляет 150 нуклеотидов. Найдите число триплетов ДНК, кодонов иРНК, антикодонов тРНК и аминокислот, соответствующих данному фрагменту. Известно, что аденин составляет 20% в данном фрагменте (двухцепочечной молекуле ДНК), найдите содержание в процентах остальных нуклеотидов.

Теперь мы украсили теорию практикой. Что может быть лучше при изучении новой темы? 🙂
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Генетический код. Биосинтез белка
теория по биологии 🌿 основы генетики
Генетическая информация и генетический код
Каждый вид имеет свой собственный, отличный от других видов, набор белков. Интересно то, что белки, выполняющие идентичные функции у разных видов могут быть похожими или даже абсолютно идентичными.
У белков есть несколько состояний их структур:
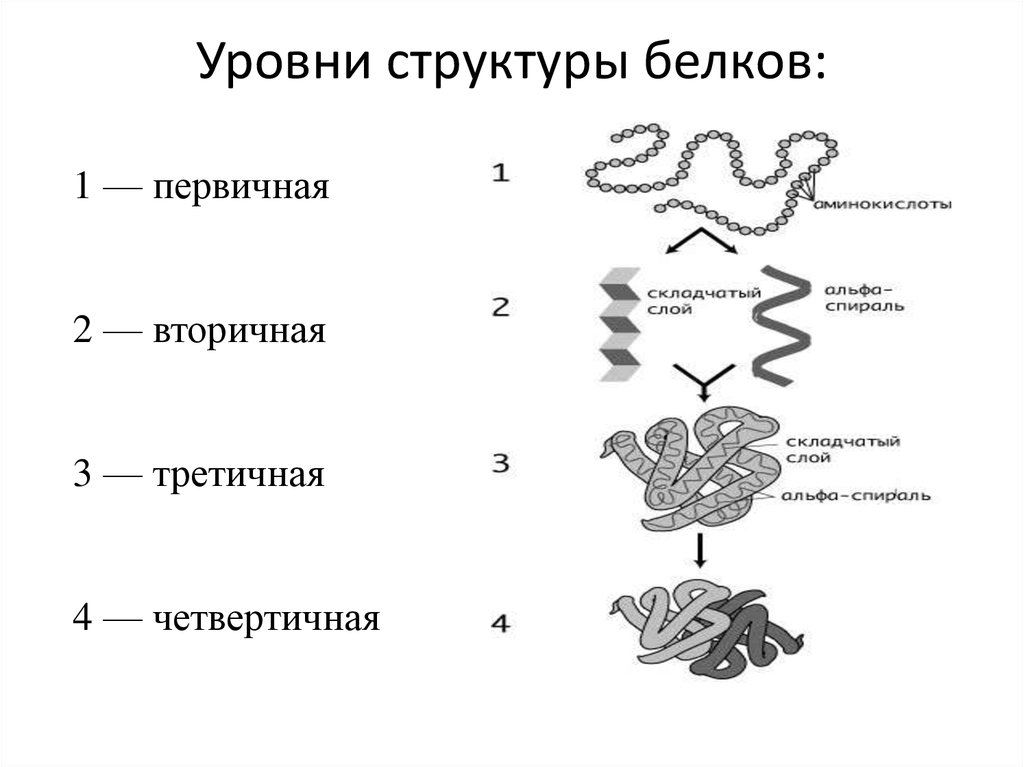
Именно первичная структура является определяющей свойства белка. Эта структура – цепь из аминокислот. Аминокислоты, в свою очередь, представляют собой ряд триплетов из нуклеотидов. Решая генетические задачи, обращаются как раз-таки к знакомой таблице:
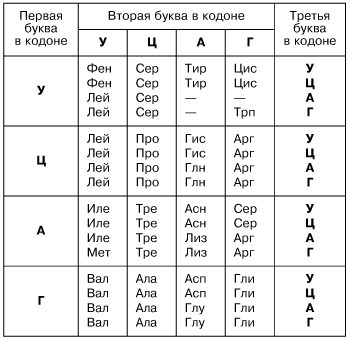
Каждая аминокислота кодируется тремя нуклеотидами, которые составляют триплет или иначе кодон. Именно последовательность нуклеотидов называется генетической информацией, а участок последовательности, в котором хранится информация о первичной структуре белка это и есть ген.
Нуклеотиды, составляющие ДНК и РНК различаются:
В состав ДНК входят:
В состав РНК входят:
Кроме того, в составе РНК (рибонуклеиновой кислоты) сахар рибоза, а ДНК (дезоксирибонуклеиновой кислоты) — дезоксирибоза. РНК — одноцепочечная, а ДНК — двухцепочечная.
Между нуклеотидами есть водородные связи. Они могут быть как двойные, так и тройные. Нуклеотиды не могу быть связаны в случайном порядке. Для этого существует принцип комплементарности ДНК, по которому аденин одной цепи ДНК соединяется с тимином другой цепи ДНК, другая пара в ДНК – гуанин – цитозин. В РНК все аналогично, за исключением того, что вместо тимина там урацил. Между парами А-Т/А-У – две водородных связи, а между парами Ц-Г – три. На письме это обозначается чёрточками: двойная связь как знак «равно», а тройная – три горизонтальные черты.
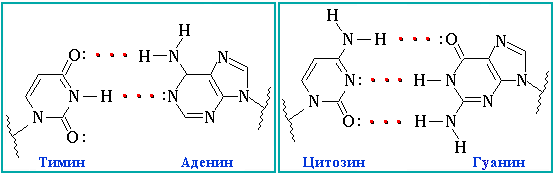
Свойства генетического кода
Транскрипция и трансляция
Из цитологии известно, что генетическая информация у эукариотических клеток заключена в ядре в виде ДНК. Однако процесс биосинтеза белка происходит в цитоплазме на рибосомах.
Спиральная цепь ДНК при раскручивается, в это время по одной из цепочек ДНК строится комплементарная цепь. Из ядра в цитоплазму информация выходит в виде информационной РНК (иРНК). иРНК комплементарная одной из цепей ДНК. Этот процесс переписывания называется транскрипцией. Полученная цепь практически идентичная другой цепи ДНК, за исключением того, что вместо тимина там урацил.В процессе участвует специальный фермент РНК-полимераза.
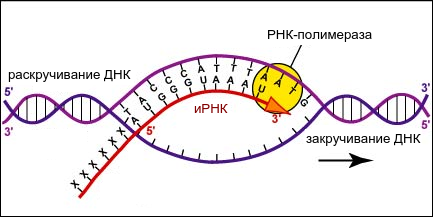
Теперь в ядре есть цепочка, которая уже начала процесс биосинтеза. Как говорилось выше, процесс ассимиляции идет на рибосомах. иРНК выходит в цитоплазму через поры ядерной мембраны
тРНК по форме напоминает лист клевера, а по принципу работы – штамп. На него, прямо как чернила, наслаиваются кодоны.
В цитоплазме начинается процесс трансляции, то есть перевод последовательности нуклеотидов информационной РНК в последовательность аминокислот белка.
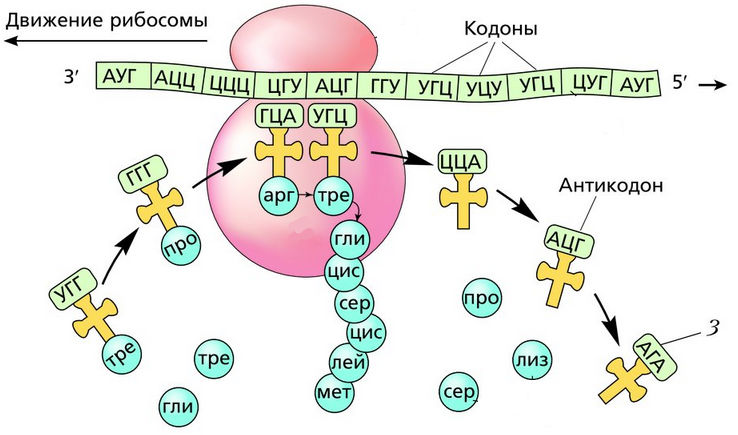
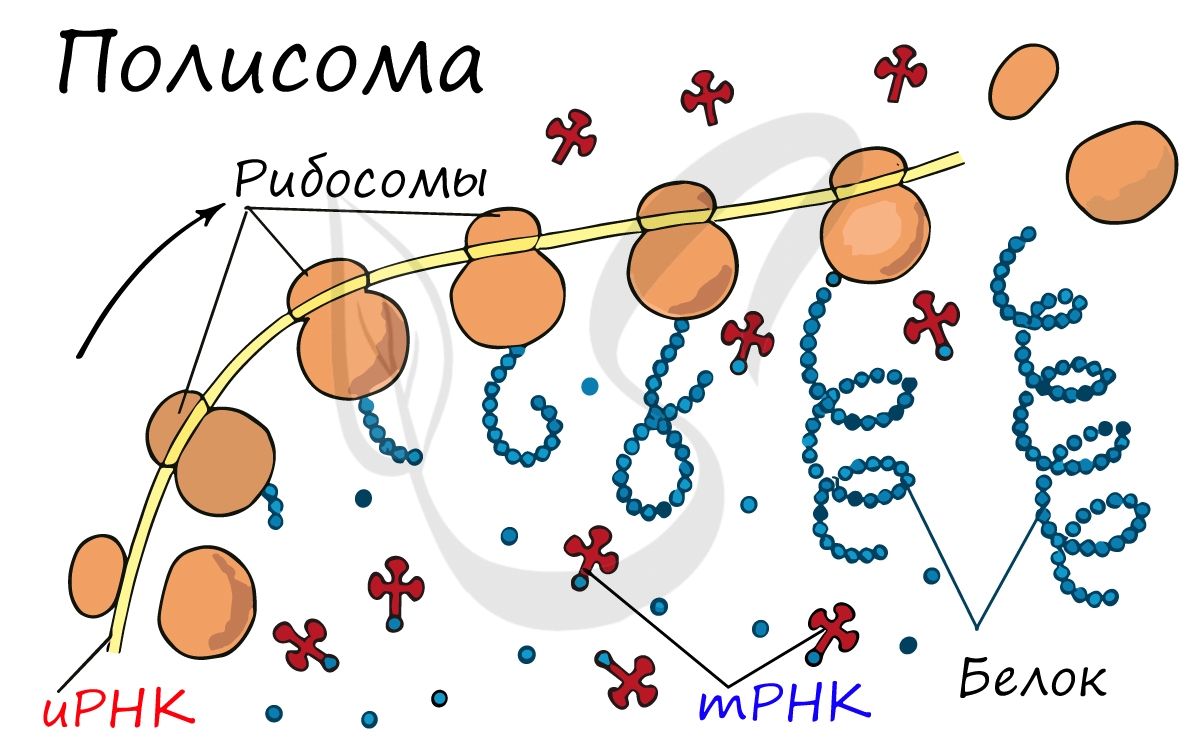
Рибосома захватывает стартовый конец цепи иРНК. Затем она начинает двигаться по цепи, одна остановка рибосомы происходит на 6-ти нуклеотидах. В это время молекула тРНК, на которых есть триплет аминокислоты «подлетает» к цепи, в месте, где находится рибосома. За время остановки рибосомы транспортная РНК успевает распознать свою пару на цепи иРНК, которая называется антикодоном. Тогда тРНК «ставит свой штамп», оставляя на цепи свой кодон. Между нуклеотидами образуются водородные связи. Так нарастает новая цепь. На одной информационной РНК работает сразу много рибосом, поэтому работа идет очень быстро. Совокупность рибосом, синтезирующих на одной иРНК, называется полисомой.
По окончанию процесса биосинтеза, цепочка отсоединяется от рибосомы и принимает свою природную структуру: вторичную, третичную или четвертичную.
pазбирался: Надежда | обсудить разбор | оценить
В современной генетической инженерии часто применняют технологии, связанные с гомологичной рекомбинацией ДНК непосредственно в живом объекте. Один из примеров – система CRE-Lox P. Lox P – это последовательность нуклеотидов в ДНК фага Р1. Она состоит из 34 нуклеотидов. В середине располагается несимметричная последовательность из 8 нуклеотидов (показана серой стрелкой на рисунке). По краям располагаются так называемые палиндромные последовательности из 13 нуклеотидов (выделены на рисунке как пунктирные блоки). Они симметричны (чтобы в этом убедиться, достаточно прочитать обе последовательность от 5´- конца к 3´- концу). Именно эти палиндромные участки узнаёт особый фермент, вызывающий рекомбинацию, который обозначают CRE. Будем в дальнейшем называть этот фермент рекомбиназой CRE. Для того, чтобы состоялась рекомбинация, два сайта Lox P должны расположиться параллельно друг другу. Рекомбиназа CRE узнает эти сайты, внесет в ДНК разрезы в определённых местах, а затем 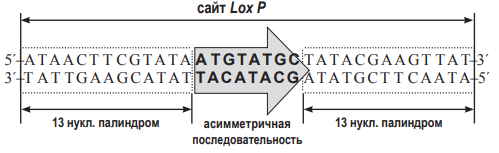 соединит по-новому две нити ДНК (т.е. произойдет рекомбинация). Аналогично работает и другая система гомологичной рекомбинации – Flp-FRT, обнаруженная у пекарских дрожжей. Сайт FRT – это последовательность ДНК, которую узнает свой фермент гомологичной рекомбинации – флиппаза (Flp).
соединит по-новому две нити ДНК (т.е. произойдет рекомбинация). Аналогично работает и другая система гомологичной рекомбинации – Flp-FRT, обнаруженная у пекарских дрожжей. Сайт FRT – это последовательность ДНК, которую узнает свой фермент гомологичной рекомбинации – флиппаза (Flp). 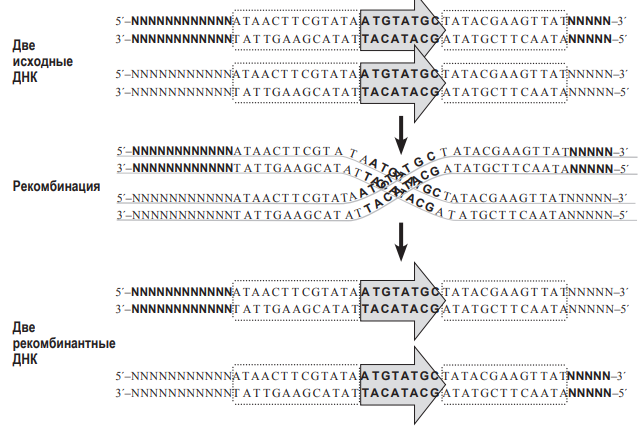 При рекомбинации две молекулы ДНК должны ориентироваться параллельно друг другу сайтами FRT, и только в этом случае произойдёт рекомбинация. Заметим, что флиппаза Flp узнает только свою последовательность FRT, но не может работать с сайтами Lox P, а рекомбиназа CRE узнает только свои сайты Lox P, но не работает с сайтами FRT. Предварительное доказательство (лемма) к задаче 9 (5 баллов). 1. Докажем, что при гомологичной рекомбинаци по «перевёрнутым» (инвертированным) повторам происходит «переворот» последовательности ДНК, находящейся между повторами. Для этого нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек.
При рекомбинации две молекулы ДНК должны ориентироваться параллельно друг другу сайтами FRT, и только в этом случае произойдёт рекомбинация. Заметим, что флиппаза Flp узнает только свою последовательность FRT, но не может работать с сайтами Lox P, а рекомбиназа CRE узнает только свои сайты Lox P, но не работает с сайтами FRT. Предварительное доказательство (лемма) к задаче 9 (5 баллов). 1. Докажем, что при гомологичной рекомбинаци по «перевёрнутым» (инвертированным) повторам происходит «переворот» последовательности ДНК, находящейся между повторами. Для этого нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек.  Затем «изогнём» молекулу так, чтобы повторы, обозначенные стрелками, встали параллельно друг другу. После обмена участками и «распрамления» окажется, что центральная часть между повторами «перевернулась».
Затем «изогнём» молекулу так, чтобы повторы, обозначенные стрелками, встали параллельно друг другу. После обмена участками и «распрамления» окажется, что центральная часть между повторами «перевернулась». 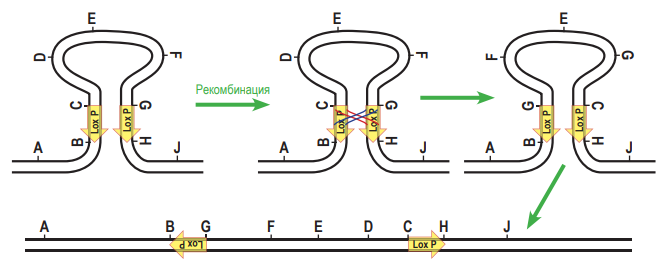 2. Докажем, что при гомологичной рекомбинаци по прямым повторам происходит образование кольцевой ДНК, при этом из линейной последовательности ДНК «удаляется» участок, находящейся между повторами. Для этого используем тот же приём: нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек. Только в этом случае для того, чтобы прямые повторы встали параллельно друг другу, придётся хитроумно изогнуть молекулу так, чтобы от конца одного из повторов (точка С) шли точки D, E, F, а потом начинался новый повтор (в точке G). После рекомбинации точки С и G поменяются местами, и в результате получится кольцевая ДНК (C, D, E, F, G) и линейный участок (A, B, H, J). Будем считать, что кольцевая ДНК как бы «исчезает» (не может реплицироваться в клетке).
2. Докажем, что при гомологичной рекомбинаци по прямым повторам происходит образование кольцевой ДНК, при этом из линейной последовательности ДНК «удаляется» участок, находящейся между повторами. Для этого используем тот же приём: нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек. Только в этом случае для того, чтобы прямые повторы встали параллельно друг другу, придётся хитроумно изогнуть молекулу так, чтобы от конца одного из повторов (точка С) шли точки D, E, F, а потом начинался новый повтор (в точке G). После рекомбинации точки С и G поменяются местами, и в результате получится кольцевая ДНК (C, D, E, F, G) и линейный участок (A, B, H, J). Будем считать, что кольцевая ДНК как бы «исчезает» (не может реплицироваться в клетке). 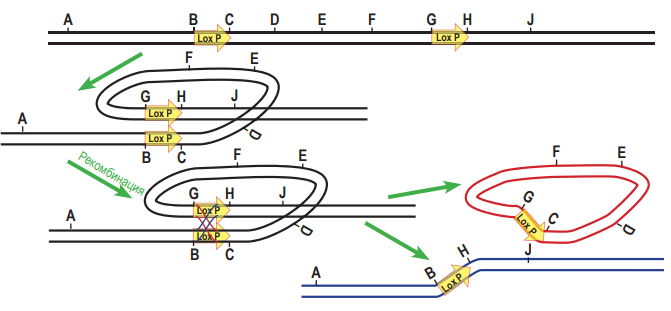 А. Поскольку после 35S-промотора на той же цепи ДНК располагается кодирующая часть гена DsRed, клетки должна светиться красным светом. Б. Рекомбиаза CRE узнаёт последовательнсоти LoxP. Если повторы расположены инвертированно, то произойдёт «переворот» последовательности ДНК, расположенной между повторами. Таким образом, после рекомбинации конструкция будет выглядеть следущим образом:
А. Поскольку после 35S-промотора на той же цепи ДНК располагается кодирующая часть гена DsRed, клетки должна светиться красным светом. Б. Рекомбиаза CRE узнаёт последовательнсоти LoxP. Если повторы расположены инвертированно, то произойдёт «переворот» последовательности ДНК, расположенной между повторами. Таким образом, после рекомбинации конструкция будет выглядеть следущим образом: 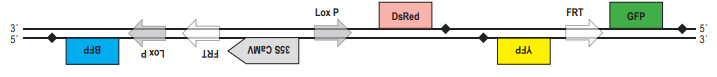 Свечение клеток изменится, поскольку после промотора на той же цепи ДНК окажется гена BFP, обестпечивающий синее свечение клеток. В. При рекомбинации по прямым повторам происходит потеря участка ДНК, расположенного между ними. Из двух повторов остаётся только один. Таким образом, после рекомбинации по сайтам FRT конструкция будет выглядеть следующим образом:
Свечение клеток изменится, поскольку после промотора на той же цепи ДНК окажется гена BFP, обестпечивающий синее свечение клеток. В. При рекомбинации по прямым повторам происходит потеря участка ДНК, расположенного между ними. Из двух повторов остаётся только один. Таким образом, после рекомбинации по сайтам FRT конструкция будет выглядеть следующим образом: 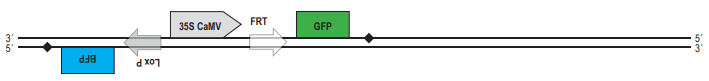 Клетки будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP. Г. После действия рекомбиназы CRE те последовательности, на которые может действовать флип паза Flp, «перевернулись», и вместо прямых стали инвертрованными. После рекомбинации участок между ними также должен «перевернуться»:
Клетки будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP. Г. После действия рекомбиназы CRE те последовательности, на которые может действовать флип паза Flp, «перевернулись», и вместо прямых стали инвертрованными. После рекомбинации участок между ними также должен «перевернуться»: 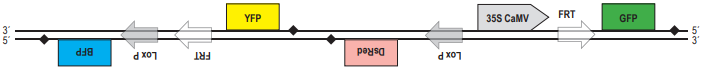 В этом случае клетки также будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP.
В этом случае клетки также будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP.
pазбирался: Надежда | обсудить разбор | оценить
pазбирался: Надежда | обсудить разбор | оценить
Сначала найдём место расщепления плазмиды рестриктазой BglII:  Таких участков оказывается два. В результате расщепления из плазмиды выщепляется короткий фрагмент:
Таких участков оказывается два. В результате расщепления из плазмиды выщепляется короткий фрагмент:  Остаётся укороченная линейная ДНК, содержащая интактный ген устойчивости к ампицилину и расщеплённый ген устойчивости к эритромицину.
Остаётся укороченная линейная ДНК, содержащая интактный ген устойчивости к ампицилину и расщеплённый ген устойчивости к эритромицину.  При сшивании липких концов ДНК-лигазой наиболее часто будут соединяться концы этой молекулы и образовываться кольцо длиной 4163 нуклеотида. Такая ДНК будет сообщать клеткам устойчивость к ампицилину и не даст устойчивости к эритромицину. Второй фрагмент из-за небольшой длины не может замкнуться в кольцо. Второй вариант лигирования приводит к сшиванию липких концов двух фрагментов. Он происходит примерно в 10 раз реже, а после сшивки вторая пара липких концов скорее всего также, как и исходный фрагмент замкнётся в кольцо. Таких колец из пары фрагментов может образоваться 4 вида: димеры большого фрагмента в двух разных ориентациях (правый конец с левым концом второго фрагмента и левый конец с правым концом второго фрагмента или правый с правым и левый с левым) и соединения большого и малого фрагмента в двух разных ориентациях (вариант исходной плазмиды и инверсия малого фрагмента). Из них только в варианте исходной плазмиды восстанавливается устойчивость к эритромицину. Линейная молекула, образованная сшиванием двух фрагментов, может присоединить ещё один фрагмент с ещё в 10 раз меньшей частотой. Такие фрагменты в дальнейшем будут циклизоваться в плазмиды трёх размеров: из трёх больших фрагментов, из двух больших и одного малого и одного большого и двух малых. Три малых фрагмента дадут короткую последовательность, которая не сможет замкнуться в кольцо и существовать в клетке. В каждом размерном классе будет несколько вариантов с разной ориентацией фрагментов. Только в одном из них восстановится ген устойчивости к эритромицину: правый конец большого фрагмента соединяется с левым концом малого фрагмента, а правый конец малого фрагмента – с левым концом второго большого фрагмента, а оставшиеся концы двух больших фрагментов соединяются с образованием кольцевой плазмиды длиной 8363 пары нуклеотидов. Доля таких молекул будет менее 1% всех плазмид. Вероятность образования плазмид из 4 и более фрагментов ещё на порядок ниже и их обнаружение при данном числе полученных трансформированных клеток нереально. А. Так как расщепление рестриктазой не затрагивает ген устойчивости к ампицилину, все клетки, в результате трансформации получившие любую плазмиду, будут устойчивы к ампицилину и вырастут на среде с этим антибиотиком. Таким образом из 33506 выросших колоний плазмиду получили 578, выросших на ампицилине. Эффективность трансформации представляет долю трансформированных клеток от общего их числа, т.е. 573 : 51366 × 100% = 1.12% Б. На эритромицине могут вырасти только те клетки, в которые попали плазмиды, в которых в результате лигирования восстановится последовательность нуклеотидов в гене устойчивости к этому антибиотику, расщеплённому рестриктазой. Остальные плазмиды, полученные по приведённой методике, будут содержать либо ген с выщепленным коротким фрагментом, что приведёт либо к утрате стартового кодона (если обозначенный зелёным цветом кодон является стартовым), либо к сдвигу рамки считывания (т.к. число удалённых нуклеотидов не кратно трём), либо, при инверсии короткого фрагмента, к появлению стоп-кодонов т.е. прекращению синтеза белка. Таким образом большинство полученных плазмид не обеспечат устойчивости к эритромицину. В. Рост на эритромицине могут обеспечить только плазмиды, несущие восстановленную последовательность гена устойчивости. Такие плазмиды могли образоваться из одного большого и одного малого фрагмента (4200 пар, исходная плазмида)) или из двух больших и одного малого (8363 пары, начало и конец гена из разных копий большого фрагмента). Г. Получается 1 размер из одного большого фрагмента, два размерных класса из двух фрагментов и три размерных класса из трёх фрагментов, то есть 6 размерных классов. (В реальности различить по длине плазмиды, отличающиеся на длину малого фрагмента, т.е. менее чем на 0,5%, невозможно. Поэтому в эксперименте, например на электрофореграмме, будут видны лишь три размерных класса, соответствующие 1, 2 или 3 копиям большого фрагмента.)
При сшивании липких концов ДНК-лигазой наиболее часто будут соединяться концы этой молекулы и образовываться кольцо длиной 4163 нуклеотида. Такая ДНК будет сообщать клеткам устойчивость к ампицилину и не даст устойчивости к эритромицину. Второй фрагмент из-за небольшой длины не может замкнуться в кольцо. Второй вариант лигирования приводит к сшиванию липких концов двух фрагментов. Он происходит примерно в 10 раз реже, а после сшивки вторая пара липких концов скорее всего также, как и исходный фрагмент замкнётся в кольцо. Таких колец из пары фрагментов может образоваться 4 вида: димеры большого фрагмента в двух разных ориентациях (правый конец с левым концом второго фрагмента и левый конец с правым концом второго фрагмента или правый с правым и левый с левым) и соединения большого и малого фрагмента в двух разных ориентациях (вариант исходной плазмиды и инверсия малого фрагмента). Из них только в варианте исходной плазмиды восстанавливается устойчивость к эритромицину. Линейная молекула, образованная сшиванием двух фрагментов, может присоединить ещё один фрагмент с ещё в 10 раз меньшей частотой. Такие фрагменты в дальнейшем будут циклизоваться в плазмиды трёх размеров: из трёх больших фрагментов, из двух больших и одного малого и одного большого и двух малых. Три малых фрагмента дадут короткую последовательность, которая не сможет замкнуться в кольцо и существовать в клетке. В каждом размерном классе будет несколько вариантов с разной ориентацией фрагментов. Только в одном из них восстановится ген устойчивости к эритромицину: правый конец большого фрагмента соединяется с левым концом малого фрагмента, а правый конец малого фрагмента – с левым концом второго большого фрагмента, а оставшиеся концы двух больших фрагментов соединяются с образованием кольцевой плазмиды длиной 8363 пары нуклеотидов. Доля таких молекул будет менее 1% всех плазмид. Вероятность образования плазмид из 4 и более фрагментов ещё на порядок ниже и их обнаружение при данном числе полученных трансформированных клеток нереально. А. Так как расщепление рестриктазой не затрагивает ген устойчивости к ампицилину, все клетки, в результате трансформации получившие любую плазмиду, будут устойчивы к ампицилину и вырастут на среде с этим антибиотиком. Таким образом из 33506 выросших колоний плазмиду получили 578, выросших на ампицилине. Эффективность трансформации представляет долю трансформированных клеток от общего их числа, т.е. 573 : 51366 × 100% = 1.12% Б. На эритромицине могут вырасти только те клетки, в которые попали плазмиды, в которых в результате лигирования восстановится последовательность нуклеотидов в гене устойчивости к этому антибиотику, расщеплённому рестриктазой. Остальные плазмиды, полученные по приведённой методике, будут содержать либо ген с выщепленным коротким фрагментом, что приведёт либо к утрате стартового кодона (если обозначенный зелёным цветом кодон является стартовым), либо к сдвигу рамки считывания (т.к. число удалённых нуклеотидов не кратно трём), либо, при инверсии короткого фрагмента, к появлению стоп-кодонов т.е. прекращению синтеза белка. Таким образом большинство полученных плазмид не обеспечат устойчивости к эритромицину. В. Рост на эритромицине могут обеспечить только плазмиды, несущие восстановленную последовательность гена устойчивости. Такие плазмиды могли образоваться из одного большого и одного малого фрагмента (4200 пар, исходная плазмида)) или из двух больших и одного малого (8363 пары, начало и конец гена из разных копий большого фрагмента). Г. Получается 1 размер из одного большого фрагмента, два размерных класса из двух фрагментов и три размерных класса из трёх фрагментов, то есть 6 размерных классов. (В реальности различить по длине плазмиды, отличающиеся на длину малого фрагмента, т.е. менее чем на 0,5%, невозможно. Поэтому в эксперименте, например на электрофореграмме, будут видны лишь три размерных класса, соответствующие 1, 2 или 3 копиям большого фрагмента.)
pазбирался: Надежда | обсудить разбор | оценить
По принципу комплементарности строим



