С каким белком непосредственно связываются ионы кальция
Ионы Са2+ представляют собой вторичный мессенджер, принимающий участие в многочисленных процессах передачи сигнала в различных клетках. У высших организмов внутриклеточный Са2+ участвует в таких разнообразных процессах, как синаптическая передача, мышечное сокращение, секреция инсулина, оплодотворение и экспрессия генов. В данной статье мы рассмотрим вопросы регуляции мышечного сокращения и сердечного ритма ионами Са2+.
В начале процесса сопряжения возбуждения и сокращения ионы Са2+ начинают поступать в цитозоль, а по мере возвращения клетки в состояние покоя выходят оттуда. Такое увеличение и снижение концентрации Са2+ в цитозоле представляют собой фазный момент в процессе сопряжения возбуждения и сокращения. Для его протекания необходимо участие нескольких типов Са2+-транспортных белков.
В клетках сердечной мышцы процесс возбуждения и сокращения подразделяется на четыре стадии.
На первой стадии, при деполяризации плазматической мембраны (сарколеммы) на ней генерируется электрический сигнал. При этом, благодаря входящему потенциалу действия, мембранный потенциал приобретает более положительное значение по сравнению с потенциалом покоя. Потенциал-зависимые Са2+-каналы (которые называются Cav1.2 Са2+-каналы) улавливают эти изменения мембранного потенциала и открываются (фаза 2 потенциала действия клеток миокарда). Небольшой поток ионов Са2+ начинает транспортироваться в клетку в направлении электрохимического градиента.
На второй стадии при поступлении Са2+ через Cav1.2 Са2+-каналы начинается высвобождение Са2+ из саркоплазматического ретикулума, в котором он депонирован в миллимолярных концентрациях. Выход Са2+ из саркоплазматического ретикулума происходит через особые каналы, называемые рианодиновые рецепторы (RyRs). В клетках сердечной мышцы этот процесс носит название Са2+-зависимый выход Са2+. Количество Са2+, выходящее в цитозоль из саркоплазматического ретикулума, в несколько раз превышает поступающее в цитозоль через сарколемму.
Са2+-зависимые Са2+-каналы саркоплазматического ретикулума клеток миокарда называются рианодиновыми рецепторами, поскольку они специфически связывают растительный алкалоид рианодин, блокирующий их действие. В различных клетках экспрессируются разные каналы внутриклеточного высвобождения Са2+, которые открываются в ответ на разнообразные сигналы, вызывающие мышечное сокращение. Основной вид каналов, высвобождающих Са2+ из саркоплазматического ретикулума клеток миокарда, относится к изоформе RyR2.
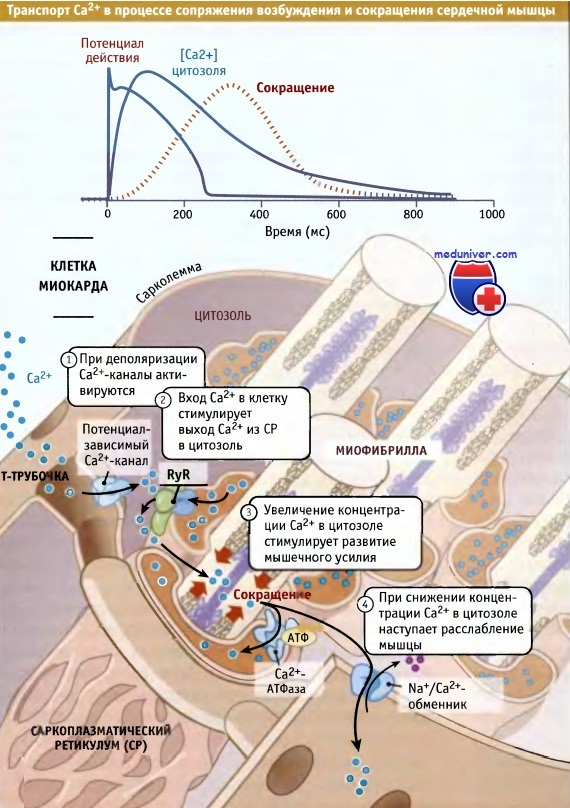 При генерации и распространении потенциала действия в клетках миокарда концентрация Са2+ в цитозоле увеличивается и снижается.
При генерации и распространении потенциала действия в клетках миокарда концентрация Са2+ в цитозоле увеличивается и снижается.
Это обеспечивается несколькими различными типами белков, транспортирующих Са2+.
На третьей стадии увеличение концентрации ионов Са2+ в цитозоле приводит к активации Са2+-зависимого белка тропонина С, который стимулирует сокращение мышечных волокон. Для эффективной активации всех внутриклеточных микрофиламентов и сокращения сердечной мышцы необходимо увеличение концентрации Са2+ в цитозоле со 100 нМ до 1,0 мкМ.
На четвертой стадии, когда Са2+ вытесняется из цитозоля, наступает расслабление мышцы. Вытеснение Са2+ происходит по нескольким механизмам. Основной из них — мобилизация Са2+ обратно в депо саркоплазматического ретикулума с помощью Са2+-АТФазы. Этот фермент осуществляет функцию насоса и направляет обратно в депо Са2+, который высвободился из саркоплазматического ретикулума при участии RyRs. Наряду с этим, Са2+ удаляется из цитозоля при участии таких Са2+- транспортных белков, как Na+/Са2+-обменник плазматической мембраны.
Этот обменник выводит из цитозоля небольшую часть Са2+, поступившего через потенциал-зависимые Cav1.2 Са2+-каналы. Немного Са2+ также обменивается между цитозолем и митохондриями.
В общем, за небольшими исключениями, процессы возбуждения и сокращения в скелетных и сердечной мышцах сходны. В отличие от клеток миокарда, потенциал-зависимые Са2+-каналы плазматической мембраны скелетных мышц представлены другой изоформой и стимулируют выход Са2+ из саркоплазматического ретикулума при физическом взаимодействии с изоформой рианодинового рецептора — RyRl. Наряду с этим, активация скелетных мышц может постепенно нарастать за счет вовлечения в процесс все большего количества мышечных волокон, что приводит к увеличению силы сокращения.
Таким образом, активация скелетных мышц может варьировать от коротких одиночных до повторяющихся тетанических сокращений и в конце концов ограничивается степенью мышечной усталости.
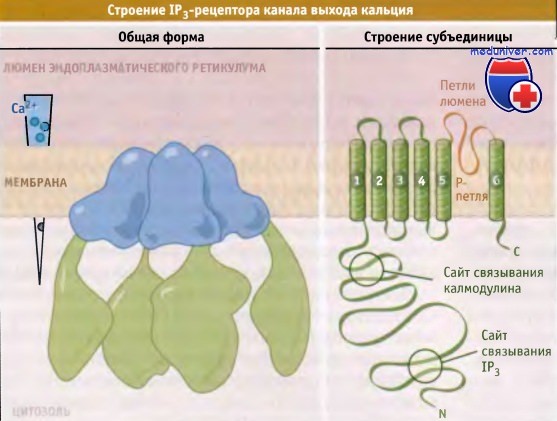 Предполагаемая структура инозитол-1,4,5-трифосфатного рецептора эндоплазматического ретикулума.
Предполагаемая структура инозитол-1,4,5-трифосфатного рецептора эндоплазматического ретикулума.
Слева показан Са2+-градиент, создающийся по сторонам мембраны эндоплазматического ретикулума в покоящихся клетках животных.
Каналы высвобождения внутриклеточного Са2+ относятся к числу уникальных ионных каналов. Они подразделяются на две группы: RyRs включаются и выключают ся под действием Са2+ или при прямом взаимодействии с Са2+-каналами плазматической мембраны, и близкие к ним рецепторы инозитол-1,4,5-трифосфата (IP3Rs), которые регулируются IР3. Четыре RyR или IP3R субъединицы собираются в симметрично организованный комплекс, образующий канал. Оба типа каналов состоят из двух доменов: порового и большого цитоплазматического, снабженного воротным механизмом. Предполагается, что канал IP3Rs имеет шесть трансмембранных сегментов и одну поровую петлю на субъединицу. Близкая структура постулируется и для RyRs.
Са2+-каналы RyR представляют собой самые большие из известных ионных каналов. Структура их доменной организации получена при трехмерном моделировании с использованием данных электронно-микроскопических исследований. Эти каналы в 10 раз больше, чем Na+-, Са2+- или К+-каналы. Каждая субъединица RyR состоит примерно из 5000 остатков аминокислот, почти вдвое превышая размер частично гомологичной IP3R субъединицы. Величина поровых доменов RyRs и IP3Rs примерно такая же, как для К+-каналов. Большие цитоплазматические домены RyRs и IP3Rs контролируют открытие-закрытие каналов с помощью Са2+ и IР3, подобно тому, как это имеет место в лиганд-зависимых К+-каналах.
Мутации, нарушающие процессы открывания-закрывания каналов внутриклеточного высвобождения Са2+, приводят к развитию различных заболеваний. Например, миссенс-мутации в гене, кодирующем белок RyR2, связаны с двумя генетическими формами аритмии и внезапной смерти при физической нагрузке. Мутантная форма RyR Са2+-канала обладает пониженным сродством к калстабину 2 (который также называется FKBP12.6). Калстабин2 представляет собой субъединицу Са2+-канала, которая стабилизирует закрытое состояние RyR миокарда, предотвращая его аберрантную активацию.
В результате мутации в белке RyR в фазе покоя или диастолы сердца, из саркоплазматического ретикулума начинается усиленный выход ионов Са2+. Более того, разрегулирование RyR2 при болезнях сердца увеличивает вероятность ухудшения сердечной функции и наступления внезапной смерти. К общему механизму, вероятно, относится внутриклеточная утечка Са2+, которая за счет аберрантной деполяризации мембраны может вызвать угрожающую аритмию.
Мутации в гене, кодирующем RyR-изоформу клеток скелетных мышц, также приводят к аберрантному выходу внутриклеточного кальция, что служит причиной заболевания, известного под названием злокачественная гипертермия. Больные крайне чувствительны к неконтролируемому внутриклеточному выходу Са2+, при этом у них поднимается температура и отмечается генерализованная мышечная контрактура. При применении в ингаляциях некоторых анестетиков и мышечных релаксантов у больных могут наступить серьезные нарушения метаболических процессов, представляющие угрозу для жизни.
Мутации в гене, кодирующем белок потенциал-зависимых Са2+-каналов плазматических мембран скелетных мышц, который физически взаимодействует с RyRs и активирует его, также обеспечивают восприимчивость к развитию злокачественной гипертермии.
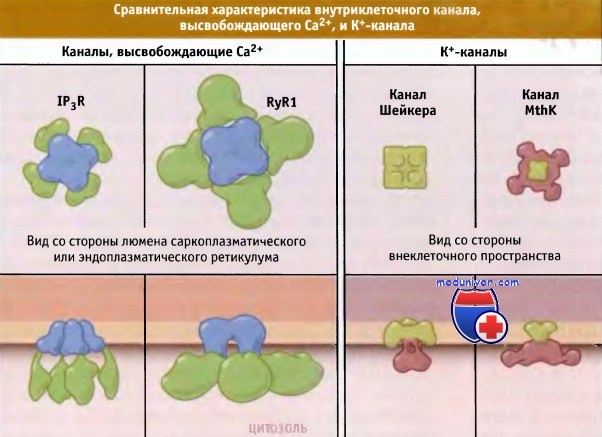 Два внутриклеточных канала, высвобождающих Са2+:
Два внутриклеточных канала, высвобождающих Са2+:
IР3-рецептор (IP3R) и рианодиновый рецептор (RyR), и два К+-канала: потенциал-зависимый канал Шейкера и Са2+-зависимый канал MthK различаются по размерам.
Поровые домены обозначены голубым и желтым цветом соответственно.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Ca++, Na+, K+ (венозная кровь) в Москве
Определение уровня кальция, натрия, калия в крови для диагностики электролитных нарушений в организме.
Приём и исследование биоматериала
Когда нужно сдавать анализ Ca++, Na+, K+?
Подробное описание исследования
Электролитный обмен — важная часть общего обмена веществ. С его помощью поддерживается постоянная среда в организме для нормальной работы клеток. Калий, натрий, кальций являются важными катионами (положительно заряженными ионами). Они участвуют в поддержании осмолярности (концентрации ионов) и кислотно-щелочного равновесия крови, регулируют метаболические процессы, влияя на работу ферментов, и обладают рядом специфических свойств. Все три макроэлемента регулируют передачу импульса в нервной и мышечной ткани, обеспечивая движение мышц и, в частности, влияют на сократимость и возбудимость сердца.
Натрий и кальций содержатся в основном вне клеток, в том числе плазме, тогда как калий преимущественно находится внутри них. Такой баланс необходим для поддержания определенного заряда мембраны клетки, с помощью которого возможна передача импульса, а также транспорт веществ в клетку и из неё (глюкозы, аминокислот).
Натрий (Na+) — основной внеклеточный катион. Поваренная соль (NaCl) является главным источником натрия. Макроэлемент поддерживает нормальное осмотическое давление в жидкостях организма, что обеспечивает их правильную работу. Так, при недостатке Na возникают симптомы обезвоживания, а при избытке — отеки.
Выводится натрий через почки и часть его вновь поступает в кровь (реабсорбируется). Благодаря воздействию гормонов на этот процесс, регулируется его уровень в плазме. Так, гормон почек ангиотензин II увеличивает реабсорбцию, а вазопрессин (гормон гипофиза) — уменьшает.
К повышению уровня натрия в сыворотке (гипернатриемии) приводит избыточное поступление его с пищей, нарушение работы почек и надпочечников, повышенные потери воды через желудочно-кишечный тракт (рвота, диарея), кожу (усиленное потоотделение). Симптомы — отеки, одышка, высокое артериальное давление.
Причины снижения макроэлемента (гипонатриемии) — малое поступление с пищей, рвота, диарея (при неправильном восполнении электролитов), заболеваниях почек, сердечной недостаточности. Проявляется гипонатриемия слабостью, потерей сознания, низким артериальным давлением, уменьшением выделения мочи, снижением эластичности кожи.
Калий (K+) поступает с пищей (фруктами, овощами) и всасывается в тонком кишечнике. Часть K+ при фильтрации почками реабсорбируется, этот процесс усиливает гормон альдостерон. На количество макроэлемента в плазме влияет pH крови — при ацидозе (увеличение кислотности) калий выходит из клеток, а при алкалозе (уменьшение кислотности) — наоборот.
Даже незначительные колебания его уровня могут привести к нарушениям работы организма. Признаками уменьшения концентрации (гипокалиемии) являются: снижение тонуса мышц, ослабление рефлексов, общая слабость. При повышении калия (гиперкалиемии) у человека может возникнуть изменение кожной чувствительности, парезы и параличи, нарушение сердечного ритма вплоть до остановки сердца.
Кальций (Ca2+) в плазме находится в трех формах: ионизированной, связанной с белками и анионами. Общий кальций плазмы — совокупность этих форм. Является основным макроэлементом для минерализации костей и зубов. Также Ca2+ участвует в передачи импульса в нервной и мышечной тканях и необходим в регуляции свертывания крови.
Макроэлемент поступает в организм преимущественно с пищей, содержится в молочных продуктах, рыбе, яйцах, зеленых овощах, орехах. В регуляции обмена Ca2+ принимают участие кальцитриол (активная форма витамина D) и паратгормон. Они увеличивают содержание кальция в крови путем усиления всасывания его в кишечнике, резорбции (разрушении) костной ткани и увеличения реабсорбции почками. Снижает уровень макроэлемента в плазме гормон кальцитонин, в основном за счет минерализации костей. Таким образом поддерживается оптимальный уровень кальция в организме.
Низкий уровень макроэлемента в сыворотке (гипокальциемия) связан с симптомами нарушения нервно-мышечного возбуждения — онемением, слабостью и болями в ногах, нарушением сердечной сократимости. Повышенная концентрация кальция (гиперкальциемия) проявляется снижением аппетита, тошнотой, болями в животе, аритмией.
Комплексный анализ крови на калий, натрий, кальций позволяет наиболее полно оценить состояние электролитного обмена. Это в свою очередь помогает выявить метаболические нарушения организма и своевременно принять меры для их устранения.
Кальций и биосинтез коллагена: систематический анализ молекулярных механизмов воздействия
Известно, что повышение уровней внеклеточного кальция стимулирует синтез/секрецию коллагена. В настоящей работе представлены результаты системно-биологического анализа кальций-зависимых белков протеома человека. Проведенный анализ указывает на перспективн
The increase of extra-cellular calcium level is known to stimulate collagen synthesis/secretion. This paper introduces results of systematic and biological analysis of calcium-dependent proteins of human proteome. This analysis indicates promising ways of increase in efficiency of calcium preparations for nutritional support of osseous metabolism.
Достаточный уровень биосинтеза коллагена является одним из важнейших показателей нормофизиологического метаболизма соединительной ткани. Фундаментальные и клинические исследования показали, что уровни кальция во внеклеточной среде стимулируют синтез/секрецию коллагена клетками внеклеточного матрикса соединительной ткани.
В эксперименте недостаточное потребление кальция негативно сказывается на состоянии биосинтеза коллагена в костной ткани [1]. Известно, в частности, что так называемые «блокаторы кальциевых каналов» ингибируют синтез коллагена I типа и его секрецию фибробластами, а аскорбат-анион противодействует эффектам блокаторов кальция [2] (рис. 1). Исследования с использованием изотопных меток показали, что блокаторы кальциевых каналов снижают преимущественно биосинтез коллагена, а не биосинтез неколлагеновых белков соединительной ткани [3].
.gif)
Показано также, что использование препаратов кальция стимулирует синтез коллагена, ускоряя заживление ран и переломов. Например, альгинат кальция улучшает заживление ран в эксперименте, повышая синтез коллагена I типа и соотношение количеств фибриллярного коллагена I типа и ретикулярного коллагена III типа. Скорость закрытия раны при использовании альгината увеличивалась по сравнению с контрольной группой [4].
В проведенном нами экспериментальном исследовании были изучены эффекты синергидной комбинации кальция с цинком, медью, марганцем, бором, магнием и витамином D (препарат Кальцемин Адванс) на модели резаной раны. Введение препарата в виде водной суспензии в течение 21 сут приводило к снижению среднего времени до полного заживления раны на 6 сут по сравнению с контролем (вода, p 2+ [11]. Рассмотрим и другие потенциальные возможности участия препаратов кальция в процессе биосинтеза коллагена, основные стадии которого представлены на рис. 5.
.gif)
Осуществляемый фибробластами или другими типами клеток соединительной ткани биосинтез коллагена включает в себя следующие стадии [12]:
Системно-биологический анализ Са-зависимых белков, которые могут воздействовать на биосинтез коллагена
Для установления Са-зависимых белков протеома, которые могут воздействовать на синтез коллагена, был проведен поиск с использованием перечисленных в табл. 2 биологических ролей. В результате был получен список из 15 белков (табл. 3).
.gif)
Для оценки вклада каждого из анализируемых 15 белков во взаимосвязь между уровнями кальция и коллагена была разработана балльная шкала оценки релевантности биологических функций белков, содержащая следующие пункты:
В табл. 3 приведены результаты оценки релевантности различных Са-зависимых белков по отношению к воздействию ионов Са 2+ на биосинтез коллагена. Очевидно, что наиболее интересными «таргетными белками» являются Са-чувствительный рецептор (CASR) и костный морфогенетический белок 4 (BMP4). Далее представлен более подробный анализ перечисленных в табл. 3 белков.
Са-чувствительный рецептор CASR
.gif) Са-чувствительный рецептор (CASR), или «сенсор кальция» [13], — рецептор, взаимодействующий с различными G-белками (Gα(q), Gα(i), G(q/11), G(i/o), G(12/13) и G(s)) [14]. Рецептор CASR (рис. 6) присутствует, в частности, в клетках паращитовидной железы, секретирующих паратиреоидный гормон (ПТГ) и в клетках эндотелия почечных канальцев. Врожденные повреждения гена CASR приводят к гиперкальциемическим или гипокальцемическим расстройствам: семейной гипокальциурической гиперкальциемии, неонатальному острому первичному гиперпаратиреозу, аутосомно-доминантной гипокальцемической гиперкальциурии [15]. Заметим, что осуществлению биологических эффектов рецептора CASR способствует активность калиевых каналов [16].
Са-чувствительный рецептор (CASR), или «сенсор кальция» [13], — рецептор, взаимодействующий с различными G-белками (Gα(q), Gα(i), G(q/11), G(i/o), G(12/13) и G(s)) [14]. Рецептор CASR (рис. 6) присутствует, в частности, в клетках паращитовидной железы, секретирующих паратиреоидный гормон (ПТГ) и в клетках эндотелия почечных канальцев. Врожденные повреждения гена CASR приводят к гиперкальциемическим или гипокальцемическим расстройствам: семейной гипокальциурической гиперкальциемии, неонатальному острому первичному гиперпаратиреозу, аутосомно-доминантной гипокальцемической гиперкальциурии [15]. Заметим, что осуществлению биологических эффектов рецептора CASR способствует активность калиевых каналов [16].
В паращитовидной железе CASR «измеряет» концентрации ионов Са 2+ в плазме крови и активирует внутриклеточные сигнальные пути, регулирующие секрецию ПТГ или фильтрацию катионов в почках. В остеоцитах CASR детектирует уровни внеклеточного кальция и активирует остеогенез. Высокие внеклеточные уровни ионов Ca 2+ (порядка 10 мМ) стимулируют экспрессию остеогенных маркеров, включая щелочную фосфатазу, костный сиалопротеин, коллаген, остеокальцин IA1, и увеличивают минерализацию кости. Кроме того, блокада рецептора CASR ингибирует клеточный ответ остеоцитов на изменения внеклеточной концентрации Ca2+ [17].
Также CASR является физиологическим регулятором роста, дифференцировки и выживания остеобластов и остеокластов [19]. Экспрессия рецептора CASR в хрящевой и костной ткани непосредственно регулирует скелетный гомеостаз и метаболизм соединительной ткани. Рецептор CASR вносит важный вклад в рост хрящевой пластинки, в т. ч. в процессы роста и дифференцировки остеобластов и остеокластов [20] (рис. 7).
На рис. 7 можно видеть, что остеобластогенез протекает посредством дифференциации мезенхимальных клеток-предшественников остеобластов, которые делятся и дифференцируются в преостеобласты, которые, в свою очередь, созревают в остеобласты. Зрелые остеобласты секретируют неминерализованный костный матрикс (остеоид), который минерализуется с образованием кости, в которой зрелые остеобласты становятся остеоцитами. Во время стимуляции остеобластов экспрессия активной R-формы рецептора CASR увеличивается в остеобластах, а экспрессия неактивной O-формы рецептора снижается, что повышает соотношение R:O и способствует остеокластогенезу. У молодых животных стимулируемый ионами кальция Са 2+ рецептор CASR индуцирует апоптоз зрелых остеокластов и предотвращает резорбцию кости, что ведет к интенсивному росту костной ткани.
.gif)
В эксперименте было показано, что активируемый ионами Ca 2+ рецептор стимулирует деление фибробластов и секрецию матриксных металлопротеиназ ММР-3, ММР-9 [21]. Повышение уровней внеклеточного кальция in vitro дозозависимо стимулирует деление фибробластов (рис. 8, контроль — 10 5 клеток, кальций — 2,2 × 10 5 клеток, p 2+ является наиболее убедительным механизмом коллаген-продуцирующего действия кальциевых препаратов.
Костный морфогенетический белок 4
Костный морфогенетический белок 4 индуцирует образование хряща и кости, участвует в развитии зубов, формировании конечностей и заживлении переломов. Интересно, что механическая нагрузка на кость способствует синтезу внеклеточного матрикса остеобластами на фоне увеличения уровней белков BMP-2/4 [22].
Повышение уровня внеклеточного кальция стимулирует увеличение числа остеобластов и ингибирует образование остеокластов, в частности, посредством воздействия на уровни экспрессии костных морфогенетических белков. Например, добавление 0,1–0,4 мМ CaCl2 к клеткам в культуре достоверно увеличивало уровни мРНК BMP-2 и BMP-4, количество синтезируемого коллагена I типа, оцененного по уровням карбокси-концевого пептида проколлагена I [23]. Повышенные уровни Ca 2+ увеличивают экспрессию генов белков морфогенеза костей (в частности, BMP-2) [24].
Взаимосвязи между активностью рецептора CASR и экспрессией морфогенетических белков остаются недостаточно изученными. С одной стороны, считается, что регулируемая секреция BMP-2 происходит в ответ на активацию рецептора CASR [25].
О других Са-зависимых белках и их воздействии на метаболизм коллагена
К другим Са-зависимым белкам, так или иначе влияющим на метаболизм коллагена, относятся матриксные металлопротеиназы (ММП) и некоторые ростовые факторы (табл. 3). Матриксные металлопротеиназы осуществляют протеолиз тройных спиралей коллагенов различных типов и играют роль в процессе заживления ран, ремоделировании соединительной ткани, деградации хряща, развитии и минерализации кости и заживлении переломов костей [26]. В структурах ММП содержатся Са-связывающие сайты, так что достаточные уровни кальция необходимы для деградации коллагена посредством ММП. Таким образом, ММП не могут непосредственно усиливать процессы биосинтеза коллагена.
Преобразующий фактор роста бета-1 (TGF-бета1) контролирует деление и дифференцировку многих типов клеток, в т. ч. фибробластов и остеобластов. TGF-бета1 стимулирует приток ионов Са 2+ в цитоплазму клетки [27] и непосредственно стимулирует синтез коллагена в остеобластах (р = 0,001), что имеет важное значение для образования костной ткани. На остеобластах в культуре одновременное добавление к среде Ca(OH)2 и TGF-бета1 значительно увеличило синтез белка вообще и синтез коллагена в частности (р = 0,048) [28].
Фактор роста соединительной ткани (CTGF) способствует делению и дифференцировке хондроцитов, обеспечивает клеточную адгезию фибробластов, осуществляет положительное регулирование процесса биосинтеза коллагена [31] посредством сигнальных путей Rac1/MLK3/JNK/AP-1 [32]. В частности, CTGF связывается с рецептором-2 факторов роста фибробластов (FGFR2) и стимулирует Са-зависимую внутриклеточную передачу сигнала [33].
Заключение
Улучшение биосинтеза коллагена — необходимое условие для восстановления структуры кости при срастании переломов, регенерации кости при постменопаузальном, лекарственном остеопорозе, а также при других нарушениях целостности кости. Нормализация биосинтеза коллагена способствует лучшему удержанию кальция в костной ткани и, следовательно, повышению минеральной плотности кости. Миллионам женщин старше сорока лет грозит перелом из-за хрупкости костей вследствие вымывания кальция. Каждые 5 минут в России происходит перелом кости. В особой группе риска 34 миллиона женщин России старше 40 лет, и 24 миллиона не знают об этом. Основная причина хрупкости костей — вымывание кальция и утрата коллагена. В отличие от других препаратов, Кальцемин Адванс содержит кальций и минералы, формирующие коллагеновую сетку, что удерживает кальций в костях. Она препятствует вымыванию кальция и сохраняет прочность костной ткани.
Системно-биологический анализ кальций-зависимых белков протеома человека показал, что ионы кальция стимулируют биосинтез коллагена посредством действия ионов кальция на Са-чувствительный рецептор CASR и костный морфогенетический белок BMP4.
Литература
* ГБОУ ВПО ИвГМА МЗ РФ, Иваново
** РСЦ Института микроэлементов ЮНЕСКО при РНИМУ им. Н. И. Пирогова, Москва
*** Клиника RHANA, Москва



