ПЦР-диагностика
Полимеразно-цепная реакция (ПЦР) – это одно из самых ярких достижений в сфере молекулярной биологии. Метод получил широчайшее распространение в разных областях науки. Благодаря очень высокой специфичности и чувствительности, метод ПЦР применяется в медицине, биологии, ветеринарии, криминалистике, санитарной службе и других отраслях деятельности человека.
Для анализа методом ПЦР можно использовать любые биологические материалы, которые содержат нуклеиновые кислоты (молекулу ДНК или РНК).
Принцип ПЦР- исследования
Кому мы обязаны появлением метода ПЦР?
Со слов американского биохимика Керри Мюллиса (Kary Mullis), идея идентифицировать живые организмы по короткому участку их генетического кода (ДНК) пришла ему в голову в 1983 году, по пути с работы домой. А в основе этой идеи, лежала работа другого американского биохимика, Артура Корнберга (Arthur Kornberg), которая в свое время не нашла отклика у научного сообщества.
Керри допустил возможность того, чтобы взять молекулу ДНК какого-либо организма, с помощью высокой температуры «распустить» ее спираль на две нити, специфическими маркерами-праймеры пометить уникальные для этого микроорганизма участки ДНК и затем, применив фермент ДНК-полимеразу, создать из двух нитей две новые молекулы ДНК. Но уже содержащие в себе меченные праймеры. И потом останется просто искать эти участки в диагностическом материале.
В итоге, корпорация CETUS, в которой работал Мюлис, выделила ему команду ученых. И в 1985 году, в издании Американского общества генетики человека, появилась публикация с теоретическим обоснованием ПЦР, как метода идентификации генетического материала живых организмов.
Как это все происходит в лаборатории

Выделение ДНК
Элонгация
Или синтез. После завершения процесса отжига, в реакционной смеси создают условия для активности полимеразы. Фермент, ориентируясь на молекулы праймеров (а не исходных нуклеиновых кислот), начинает синтез новых ниток ДНК/РНК. Которые становятся копиями исходных, искомых молекул нуклеиновых кислот.
Такой температурный цикл проводится 30 и более раз. В результате, даже при изначально небольшом количестве искомого генетического материала, в реакционной смеси накапливается значительное число «помеченных» праймерами нуклеиновых кислот (растет экспоненциально, с удвоением при каждом цикле).Обнаружить большие количества ДНК/РНК намного проще, за счет чего реализуется еще одно преимущество ПЦР – высочайшая чувствительность.
Детекция
Оценка результатов ПЦР проводится несколькими путями:
Преимущества методики ПЦР
Всего разработано более 10 разных методик амплификации, применяемых лабораториями в зависимости от исходных условий и поставленных целей.
Общим для них есть высокая чувствительность (для положительного результата достаточно 40 (!) или менее искомых копий ДНК в 1 мл образца, то есть вероятность ложноотрицательного ответа ничтожно мала. И очень высокая специфичность: вероятность ложноположительного ответа составляет менее 1%.
Но точность результатов сильно зависит от качества сбора диагностического материала, тщательного соблюдения всех технических требований к каждому этапу и качеству оборудования, расходных материалов (буфера, праймеров, раствора для отмывки и т.д.).

Области применения в медицине
В дерматовенерологии ПЦР используют для выявления венерических заболеваний: микоплазменной, хламидийной инфекций, сифилиса, генитального герпеса и др.
Инфекционисты активно используют ПЦР для диагностики туберкулеза, ВИЧ, вирусных гепатитов, герпеса, мононуклеоза, вируса Эпштейн-Барр и др.). А с помощью ПЦР в реальном времени, оценивая вирусную нагрузку, врачи могут составить мнение о динамике заболевания, отклике на лечение, что особенно актуально для пациентов с ВИЧ, принимающих терапию.
Также благодаря ПЦР врачи могут в течение нескольких дней с уверенностью идентифицировать коклюш и паракоклюш, выявить возбудителей эпидемии ОРВИ. Уточняются типы вируса гриппа, циркулирующие на определенной территории, на основании чего появляется возможность разработать эффективную вакцину для каждого сезона гриппа.
В течение суток или быстрее можно установить вид возбудителя кишечной инфекции, а значит – назначить адекватное лечение и обнаружить вероятный источник заражения.
Летом, ПЦР актуальна для диагностики заболеваний, передаваемых иксодовыми клещами: боррелиоза (болезни Лайма), клещевых энцефалитов.
Метод позволяет работать с любым биологическим материалом. Гемотрансмиссивные инфекции (сифилис, ВИЧ, гепатиты, боррелиоз) исследуются по пробе венозной крови или спинномозговой жидкости. Кожные болезни (герпес, грибки) – по соскобу с пораженного участка. Венерические и урологические – по образцу мочи, спермы, влагалищного отделяемого.
Так что в медицине, ПЦР применяется везде, где нужна высокая точность и быстрота получения результатов.
Лабораторные исследования, выполняющиеся методом ПЦР:
Разрывом каких связей сопровождается денатурация днк
Молекулярная масса ДНК определяется рядом методов, в том числе:
а) ультрацентрифугированием в градиенте CsCl (ММ от 200 000 до 10 9 );
Денатурация ДНК заключается в разрыве Н-связей и стэкинг-взаимодействий, что приводит к расплетанию и разделению цепей (без разрыва ковалентных связей) под действием температуры или рН.

Денатурация – процесс утраты молекулами нуклеиновых кислот третичной и вторичной структуры. В денатурированном состоянии молекулы временно нефункциональны, однако, способны восстанавливаться в результате ренатурации (в отличие от молекул белков). Денатурация нуклеиновых кислот чаще всего вызывается теплом. Первичная структура при денатурации не изменяется.
О степени денатурации судят по изменению интенсивности поглощения в ультрафиолете при l = 260 nm, поскольку дезэкранирование азотистых оснований в результате расплетания цепей устраняется, что вызывает увеличение степени поглощения раствором ДНК ультрафиолета указанной длины волны:

Денатурацию иначе называют плавлением, а температура плавления соответствует моменту 50%-ной денатурации молекулы. Температура плавления различается у разных ДНК.
Комплементарные цепи ДНК, разделенные при денатурации, при определенных условиях могут вновь соединиться в двойную спираль. Этот процесс называется ренатурацией. Если денатурация произошла не полностью, и хотя бы несколько оснований не утратили взаимодействия водородными связями, ренатурация протекает очень быстро. Ренатурация возможна даже при полностью разделенных цепях. В таком случае ренатурация требует точного совмещения цепей ДНК, которое может привести к реассоциации, и этот процесс медленный, к тому же зависит от концентрации цепей в растворе.
Ренатурация – процесс восстановления нативной (исходной, биологически активной) структуры молекулы нуклеиновой кислоты. Ренатурация подразумевает полное восстановление вторичной структуры нуклеиновой кислоты (комплементарное взаимодействие цепей и образование двунитевой спирали в случае, например, молекул ДНК). Чем сложнее молекула ДНК, тем больше времени требуется для ренатурации.
3.4 Вторичная структура ДНК
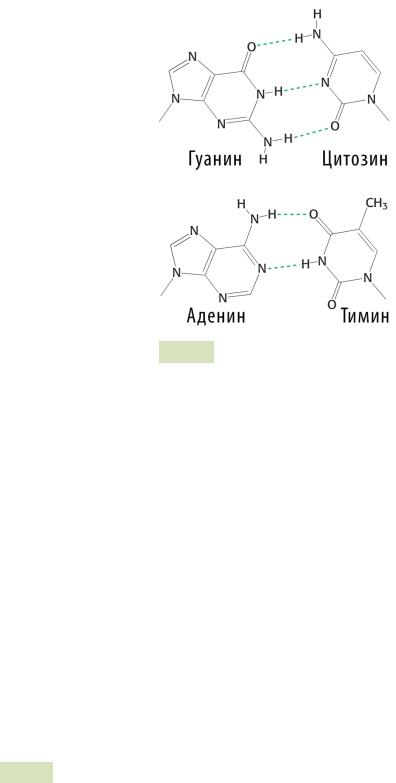
Вторичная структура ДНК представляет собой двухцепочечную спираль (см. Рис. 50 ), из двух длинных полинуклеотидных цепей, закрученных относительно общей оси. Сахарофосфатный скелет обеих цепей обращён наружу от оси спирали, а азотистые основания обращены внутрь спирали.
Две цепи удерживаются вместе с помощью водородных связей между азотистыми основаниями (аденин образует две связи с тимином, а гуанин — три связи с цитозином), гидрофобных связей, стэкинг-взаимодействий и ионных связей. Одна цепь ДНК комплементарна другой, при этом цепи антипараллельны.
Структурные особенности двойной спирали ДНК (см. Рис.
1. Две антипараллельные цепи являются правозакрученными вдоль общей оси, диаметр спирали
2. Плоскости, в которых лежат азотистые основания, образующие водородные связи, почти перпендикулярны оси спирали. В B-ДНК основания занимают центральную часть спирали, а сахарофосфатный скелет закручен и обращён наружу. Только края оснований контактируют с растворителем.
3. Каждое основание имеет приблизительно одну ширину, что придает спирали ДНК практически полную симметрию независимо от состава самих оснований. Основания в парах можно поменять местами без каких-либо последствий для спирали: Г ≡ Ц можно изменить на Ц ≡ Г или А = Т на Т = А.
Рис. 50. Вторичная структура ДНК.
Рис. 51. Вторичная структура ДНК: двойная спираль и её параметры.
4. Каноническая (идеальная) форма B- ДНК имеет 10 пар оснований в одном шаге (в одном повороте спирале). При этом на каждую пару приходится 36° (если быть точным, то число пар в шаге — 10,4, следовательно, угол составляет 34,6°). Пары оснований уло-
жены в спирали «стопками». Шаг спирали составляет 34 Å (3,4 нм).
ДНК обладает структурной гибкостью и существует в нескольких формах : A-ДНК, B-ДНК (классическая Уотсон-Криковская двойная спираль), Z-ДНК и др. A- и B-ДНК являются правозакрученными спиралями. В B-ДНК азотистые основания располагаются почти перпендикулярно к оси вращения спирали. Z-ДНК представляет собой левозакрученную спираль. Бо́льшая часть ДНК клетки находится в B-форме (см. Приложение 3).
В двойной спирали различают большую и малую бороздки — это расстояния между сахарофосфатными остовами обеих цепей в двойной спирали.
Рис. 52. Уотсон-Криковские пары комплементарных осно-
ваний: пунктиром отмечены водородные связи между атомами
Химические связи, стабилизирующие вторичную структуру ДНК:
1. Стэкинг-взаимодействия : пары оснований, уложенные в спирали «стопками» удерживаются силами Ван-дер-Ваальса. И несмотря на то, что связи между 2 парами оснований крайне слабые, их большое количество во всей молекуле ДНК является важным фактором её стабилизации.
2. Гидрофобные связи : гидрофобные азотистые основания «погружены» внутрь спирали и взаимодействуют друг с другом, это придаёт спирали дополнительную стабильность.
3. Водородные связи образуются между азотистыми основаниями цепей. Между аденином и тимином — 2 водородных связи, а между гуанином и цитозином — 3 (см. Рис. 52 ).
4. Электростатические взаимодействия : электростатическое отталкива-
ние между отрицательно заряженными остатками фосфорной кислоты дестабилизирует спираль ДНК, однако эти эффекты минимизированы присутствием ионов Mg 2+ и катионных белков (богатых лизином и аргинином).

Денатурация и ренатурация ДНК 77
3.5 Денатурация и ренатурация ДНК
ДНК в клетке термодинамически стабильна и спирализована, однако некоторые её области подвергаются «плавлению» (т.е. двойная спираль разматывается) изза происходящих процессов репликации, транскрипции, репарации и рекомбинации. Полное расплетение цепей и их разделение называют денатурацией. Денатурация происходит только in vitro (см. Рис. 53 ).
Двухцепочечная ДНК может быть денатурирована воздействием высоких температур или хаотропных агентов, таких как мочевина или гуанидин хлорид.
При медленном повышении температуры двойная спираль ДНК денатурирует постепенно, за счет нарушения стэкинг и водородных связей. Две цепи постепенно расплетаются и наконец полностью диссоциируют (разделяются). Температура, при которой половина молекулы ДНК подверглась денатурации, называется
Ренатурацией называется процесс восстановления нативной кон-
формации ДНК. Ренатурация происходит самопроизвольно при охлаждении раствора, в котором содержатся диссоциировавшие цепи ДНК.
ДНК и РНК могут образовывать гибриды по принципу комплементарности (например, матричная цепь ДНК и транскрибированная с неё пре-мРНК). Кроме того, если смешать в одном растворе денатурированные цепи ДНК различных видов, они тоже будет образовывать гибриды, но в силу того, что полностью нуклеотидные последовательности этих цепей некомплементарны (ввиду генетической изменчивости и разнообразия), такие гибриды будут считаться неполными : помимо спирализованных участков будут образовываться «вздутия», не имеющие спиральной укладки.
Степень денатурации можно измерить с помощью абсорбции ультрафиолетовых лучей. Одноцепочечные ДНК при длине волны 260 нм поглощают 12-40% света. Разумеется, области, богатые парами A = T быстрее подвергаются плавлению, чем Г ≡ Ц участки.
78 Глава 3 Нуклеиновые кислоты
3.6 Третичная структура ДНК
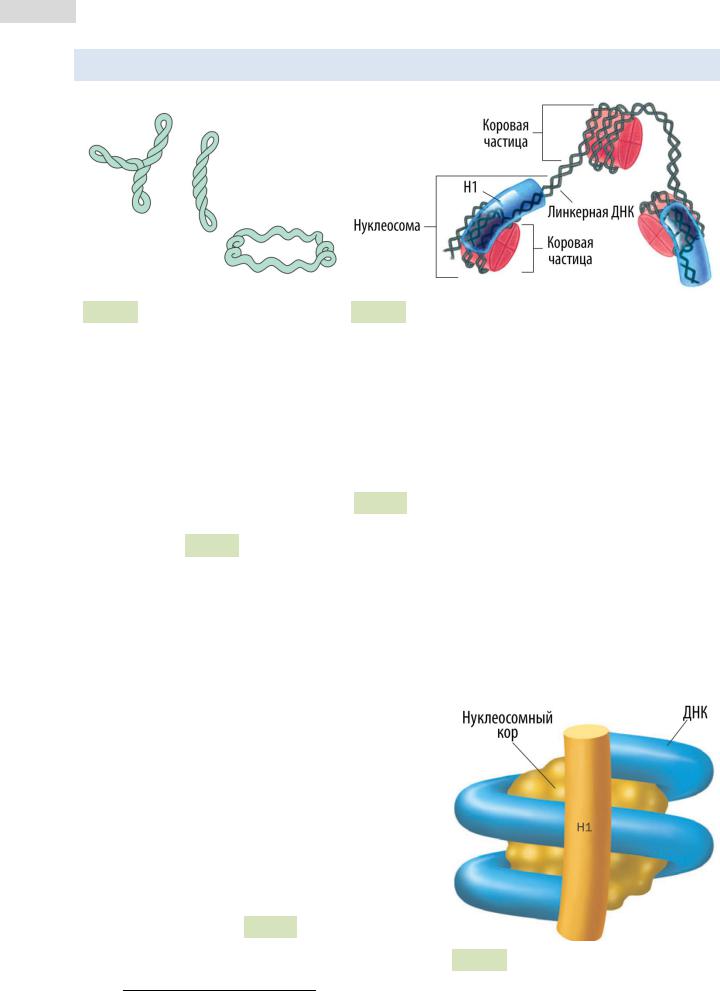
Рис. 54. Отрицательные супер- Рис. 55. Строение нуклеосомной нити витки в кольцевой бактериальной ДНК (слева) и их постепенное раскручивание (справа)
Третичная структура ДНК — это пространственная форма, которую принимает молекула ДНК по мере своего сворачивания и компактизации. Третичная структура ДНК про- и эукариот различается. Прокариотическая ДНК связана лишь с небольшим количеством белков, среди которых есть и гистоноподобные. Она обычно свёрнута в суперспираль округлой формы (см. Рис. 54 ▲ ). У эукариот степень упаковки ДНК гораздо выше, поскольку их цепи связаны с гистоновыми белками, образующими нук-
леосо́мы (см. Рис. 55 ▲ ). Нуклеосома — это комплекс нуклеосомного ко́ра и накру-
ченного на него участка ДНК. Каждый нуклеосомный кор состоит из 4 пар гисто́нов (всего 8 белковых субъединиц): H2A, H2B, H3 и H4. Гистоны — положительно заряженные белки (из-за обилия остатков аргинина и лизина), взаимодействующие с отрицательно заряженной ДНК. Комплекс гистонов и ДНК стабилизирован водород-
модействиями 1 (связь между положительно заряженным концом одной молекулы и отрицательно заряженным концом другой).
Вокруг одной нуклеосомной частицы ДНК делает 2,5 оборота (1,75 × 2). Длина участка ДНК, связанного с нуклеосомным кором, составляет 146 пар нуклеотидов. Фрагмент спирали ДНК, находящийся между двумя нуклеосомными частицами и связывающий их, называют линкерным ( линкерная ДНК ). Гистон H1 в состав нуклеосомного кора не входит, однако связан с молекулой ДНК, которая оборачивается вокруг нуклеосомного кора, в области её «вхождения» и «выхода» из кора (см. Рис. 56 ). Роль H1 заклю-
чается в поддержании плотности упаковки ДНК Рис. 56. Гистон H1 и нуклеосома.
1 См. Таблица 2 (стр. 12) и Рис. 127 (стр. 191)
Тесты из задачника 989898FF
Б. Отрицательный В. Нейтральный
в) заряд обусловлен присутствием в белке большого количества:
г) эти белки входят в состав:
д) образование этих структур способствует:
Б. Компактизации ДНК
В. Повышению отрицательного заряда ДНК Г. Транскрипции
9. Выберите один правильный ответ. Минорные основания:
А. Образуются в результате дезаминирования урацила Б. Образуют ковалентные связи, стабилизирующие 3-ю структуру РНК В. Снижают устойчивость РНК к действию нуклеаз
Г. Препятствуют спирализации определенных участков РНК
Д. Участвуют в образовании комплементарных пар
10*. Выберите один правильный ответ. Денатурация ДНК сопровождается:
А. Образованием ковалентных «сшивок» между цепями Б. Гидролизом 3′,5′-сложноэфирной связи между мономерами В. Нарушением первичной структуры цепей ДНК
Г. Разрывом водородных связей между цепями ДНК
Д. Гидролизом N-гликозидной связи в мономерах
11*. Выберите один неправильный ответ. Методом молекулярной гибридизации можно установить:
А. Различие ДНК, выделенных из организмов разных видов Б. Идентичность ДНК, выделенных из разных органов одного организма В. Видовую специфичность молекул ДНК
Г. Сходство первичной структуры ДНК, выделенной из одного и того же органа разных людей
Д. Первичную структуру ДНК
12*. Выберите один неправильный ответ. Молекулы РНК:
А. Построены из рибонуклеозидмонофосфатных остатков Б. Состоят из одной полинуклеотидной цепи В. Имеют разное строение 5′- и З’-концов Г. Содержат спирализованные участки
Д. Синтезируются в ходе репликации
13*. Выберите один неправильный ответ. Молекула мРНК:
А. Построена из нуклеозидмонофосфатов Б. Имеет полиА-последовательность на З’-конце
В. Содержит равное количество уридиловых и адениловых нуклеотидов
Г. На 5′-конце имеет «кэп» Д. Образует спирализованные участки
14. Установите соответствие.
3-А. На З’-конце имеет «кэп»
Б. Образуют с белками рибонуклеопротеиновые комплексы с разным значением S
Г. Входит в состав хроматина
2-Д. Имеет полиА-последовательность на З’-конце
15*. Выберите один неправильный ответ. Виды РНК различаются:
А. Первичной структурой Б. Молекулярной массой
В. Способом соединения нуклеотидов в полинуклеотидной цепи
16. Установите соответствие. Матрица:
Г. Метилирование ДНК
17*. Выберите один неправильный ответ. Репликация:
А. Начинается после перехода клетки в фазу синтеза Б. Предполагает образование репликативной вилки
В. Осуществляет точное воспроизведение ДНК перед каждым клеточным делением
Г. Обеспечивает многократное удвоение генома в течение S-фазы
Д. Завершается образованием тетраплоидного набора хромосом
18. Выберите наиболее полное утверждение. При репликации происходит:
А. Образование 3′,5′-фосфодиэфирных связей между мономерами Б. Локальное расхождение цепей ДНК-матрицы
В. Удвоение генома клетки.
Г. Образование фрагментов Оказаки Д. Образование нуклеотидных цепей, комплементарных нуклеотидным цепям матрицы
19*. Выберите утверждение, которое нарушает последовательность событий.
A. При участии ДНК-топоизомеразы и хеликазы образуется репликативная вилка
Б. ДНК-полимераза α начинает синтез новых цепей
B. ДНК-лигаза «сшивает» фрагменты Оказаки
Г. ДНК-полимеразы δ и ε катализируют синтез лидирующей и отстающей цепей ДНК
Д. ДНК-полимераза β вырезает праймеры и заполняет «брешь»
20*. Выберите один неправильный ответ. Репликативная вилка:
А. Представляет собой локальное расхождение цепей ДНК-матрицы Б. Перемещается вдоль спирали ДНК-матрицы В. Образуется при участии белков репликативного комплекса
Г. Необходима для одновременного синтеза двух новых цепей ДНК
Д. Поддерживается при участии ТАТА-фактора
21*. Выберите один правильный ответ. ДНК-лигаза:
А. Не входит в состав репликативного комплекса Б. Синтезирует фрагменты цепей ДНК
В. «Сшивает» фрагменты Оказаки
Г. Катализирует гидролиз 3′,5′-фосфодиэфирной связи Д. Активируется ТАТА-фактором
22*. Выберите один неправильный ответ.
Для точного воспроизведения ДНК матрицы необходимо:
А. Постепенное продвижение репликативной вилки по ДНК-матрице Б. Участие белков, обеспечивающих локальное расхождение цепей ДНКматрицы В. Доступность оснований ДНК-матрицы для комплементарного спаривания
Г. Присутствие экзонуклеазы в составе репликативного комплекса
Д. Образование праймеров с помощью ДНК-полимеразы α
23. Установите соответствие.
1. Присоединяет остатки дезоксинуклеозидмонофосфата в процессе роста лидирующей цепи ДНК
2. Синтезирует фрагмент РНК
3. Гидролизует 3′,5′-фосфодиэфирную связь в одной из цепей ДНК
2-А. ДНК-полимераза α
1-В. ДНК-полимераза δ
3-Д. ДНК-топоизомераза I
24. Выполните «цепное» задание.
а) в репликации участвует: А. ДНК-полимераза δ
Б. ДНК-инсертаза В. ПолиА-полимераза
б) фермент катализирует образование:
А. N-гликозидной связи
Б. 3′,5′-фосфодиэфирной связи
В. 5′,5′-фосфодиэфирной связи
в) связь образуется между:
А. Азотистым основанием и дезоксирибозой
Б. Последним нуклеотидом растущей лидирующей цепи и последующим
г) под действием фермента образуется цепь ДНК, которая:
А. Идентична матричной цепи
Б. Комплементарна отстающей цепи
В. Подвергается сплайсингу
д) которая синтезируется при участии:
А. Малых ядерных рибонуклеопротеинов (мяРНП)
25. Выберите один правильный ответ. Фермент теломераза:
А. Синтезирует праймер Б. Не требует затрат энергии
В. Участвует в достройке 5′-концов цепей ДНК
Г. Неактивен в быстро делящихся клетках Д. Удаляет из цепей ДНК минорные нуклеотиды
26. Установите соответствие.
А. Фермент, синтезирующий олигорибонуклеотид Б. Содержит информацию о строении регуляторного белка
3-В. Фермент, содержащий в качестве простетической группы фрагмент РНК 1-Г. Олигорибонуклеотид
2-Д. Присутствует на 5′-конце цепей ДНК
2. Теломерная последовательность
27*. Выберите один неправильный ответ. Репарация:
А. Происходит в ядре Б. Обеспечивает стабильность генома
В. Активируется в S-фазу клеточного цикла
Г. Происходит при участии ферментов эндонуклеазы и экзонуклеазы Д. Невозможна при одновременном повреждении комплементарной пары нуклеотидов
28. Выполните «цепное» задание.
а) к накоплению повреждений в ДНК приводит снижение скорости:
б) в ходе этого процесса:
А. Вырезаются дезаминированные нуклеотиды из цепей РНК
Б. Синтезируются новые цепи ДНК, идентичные матричным
В. Устраняются повреждения в ДНК
в) ферменты этого процесса обеспечивают удаление из цепей ДНК:
А. Дезаминированных нуклеотидов Б. Димеров тимина
В. Комплементарных пар поврежденных нуклеотидов
Г. Нуклеотидов, модифицированных канцерогенами Д. Продуктов депуринизации нуклеотидов
29*. Выберите один неправильный ответ. Ферменты репарации:
А. ДНК-гликозилазы разрушают N-гликозидную связь между дезоксирибозой и поврежденным основанием
Б. Эндонуклеазы гидролизуют 3′,5′-фосфодиэфирную связь только между
пиримидиновыми нуклеотидами В. Экзонуклеазы удаляют поврежденный участок цепи ДНК
Г. ДНК-полимераза β синтезирует фрагмент цепи ДНК Д. ДНК-лигаза катализирует образование 3′,5′-фосфодиэфирной связи
между вновь синтезированным и основным участками цепи ДНК
30. Выберите утверждение, которое предшествует описанной ситуации.
А Эндонуклеаза определяет место повреждения Б Экзонуклеаза «вырезает» поврежденный участок
В ДНК-полимераза β достраивает поврежденную цепь
Г. Под действием ультрафиолетовых лучей в цепи ДНК образуются пиримидиновые димеры
Д. ДНК-лигаза соединяет основной и новообразованный участки цепи
31. Выберите один правильный ответ. Апуриновые эндонуклеазы:
А. Относятся к классу лиаз
Б. Активируются в S-фазу клеточного цикла В. Образуют апуриновые сайты
Г. Отщепляют по одному нуклеотиду с З’-конца поврежденной цепи ДНК
Д. Распознают апуриновый сайт
32*. Выберите один правильный ответ. Транскрипция:
А. Происходит в S-фазу клеточного цикла Б. Всегда начинается с кодона AUG
В. Инициируется образованием праймера Г. Не требует локального расплетения двойной спирали ДНК
Д. Протекает при участии ТАТА-фактора
33*. Выберите утверждение, которое нарушает последовательность событий.
В ходе транскрипции происходит:
В. Активация фактора терминации
Г. Образование молекулы РНК, комплементарной транскриптону Д. Отделение молекулы РНК от матричной цепи ДНК
34*. Выберите один неправильный ответ. РНК-полимераза:
А. Присоединяется к промотору
Б. Раскручивает определенный участок ДНК В. Для начала синтеза не требует «затравки»
Г. Начинает синтез молекулы РНК с образования «колпачка»
Д. Для синтеза РНК использует энергию нуклеозидтрифосфатов
35*. Выберите один правильный ответ. Промотор:
А. Специфическая последовательность нуклеотидов в молекуле РНК Б. Присоединяется к репликону
В. Место присоединения РНК-полимеразы
Г. Предшествует транскриптону Д. Необратимо связывается с ТАТА-фактором
36. Установите соответствие.
1. Образуется в ядре
2. Синтезируется при участии SSB-белков
4. Не содержит антикодоновой петли
37*. Выберите один правильный ответ. Пре-мРНК:
А. Представляет собой полный транскрипт гена
Б. Последовательность триплетов, кодирующих первичную структуру белка B. На 5′-конце имеет полиА-последовательность Г.
Связывается с рибосомой в области колпачка Д. Выходит из ядра в цитоплазму
38*. Выберите один неправильный ответ.
В ходе образования зрелой мРНК происходит:
А. Разрыв 3′,5′-фосфодиэфирной связи в местах «вырезания» интронов Б. Взаимодействие пре-мРНК с мяРНП В. Образование полиА-последовательности на З’-конце мРНК
Г. Присоединение к 5′-концу мРНК «кэпа»
Д. Связывание мРНК с субъединицами рибосом
39*. Выберите один неправильный ответ. В процессе альтернативного сплайсинга:
Б. Осуществляется построение «кэпа» на 5-конце
В. Происходит гидролиз 3′,5′-фосфодиэфирной связи на границе экзонинтрон Г. мяРНП «сшивают» экзоны
Д. Образуются «зрелые» мРНК с разной первичной структурой
40. Установите соответствие.
3-А. Специфическая последовательность в составе промотора 1-Б. Участок ДНК, содержащий информацию о структуре одной полипептидной цепи
В. Концевой фрагмент цепи ДНК
2-Г. Единица репликации
Биосинтез белка (трансляция)
41*. Выберите один правильный ответ. Генетический код:
А. Порядок чередования нуклеотидов в ДНК Б. Порядок чередования нуклеотидов в РНК
В. Способ записи первичной структуры белков с помощью нуклеотидной
последовательности ДНК или РНК Г. Триплет нуклеотидов, кодирующий одну аминокислоту
Д. Набор генов, определяющий фенотипические признаки
42*. Выберите один неправильный ответ. Для генетического кода характерны:
А. Вырожденность Б. Универсальность В. Специфичность
43*. Выберите один неправильный ответ. Свойства кода:
А. Каждый кодон шифрует одну аминокислоту
Б. Каждую аминокислоту кодирует только один кодон
В. Кодоны мРНК читаются в направлении от 5′- к З’-концу Г. Одну аминокислоту могут кодировать несколько кодонов
Д. Смысл кодонов одинаков почти для всех живых организмов на Земле
44*. Выберите один правильный ответ. Коллинеарность:
А. Способ шифрования первичной структуры белков в нуклеотидной последовательности ДНК и РНК Б. Участок молекулы ДНК, содержащий информацию о первичной структуре одной полипептидной цепи
В. Триплет нуклеотидов, кодирующий включение одной аминокислоты
Г. Соответствие между последовательностью кодонов мРНК и первичной структурой белка
Д. Связь антикодона аминоацил-тРНК с кодоном мРНК
45*. Выберите один неправильный ответ.
В процессе синтеза белка принимают участие: А. Рибосомы Б. Факторы инициации
Д. Факторы элонгации
46*. Выберите один неправильный ответ.
В процессе синтеза белка на этапе инициации принимают участие: А. Субъединицы рибосом

Б. Факторы инициации В. Мет-тРНК Мет
47. Выберите один наиболее полный ответ. В ходе трансляции:
А. Участвуют факторы инициации, элонгации, терминации Б. На каждом этапе элонгации синтезируемый белок удлиняется на одну аминокислоту В. Затрачивается энергия АТФ и гуанозинтрифосфата (ГТФ)
Г. Синтезируется полипептидная цепь белка-предшественника
Д. Участвуют аминоацил-тРНК
48. Выполните «цепное» задание.
а) на рисунке изображен один из этапов в процессе синтеза белка:
б) с тРНК в пептидильном центре непосредственно связана аминокислота:
в) следующей стадией биосинтеза белка будет:
А. Образование пептидной связи Б. Элонгация В. Транслокация
Г. Связывание аминоацил-тРНК
А. Образование пептидной связи
Б. Терминация В. Транслокация
Г. Связывание аминоацил-тРНК
д) после включения в А-центр рибосомы кодона UAG наступает:
А. Элонгация Б. Инициация
49*. Выберите один правильный ответ. На каждой стации элонгации происходит:
А. Удлинение растущей пептидной цепи на одну аминокислоту
Б. Включение Мет-тРНК Мет в Р-центр В. Взаимодействие аминокислот с тРНК Г. Использование энергии АТФ Д. Освобождение готового белка
50*. Выберите один правильный ответ. Антикодон:
А. Триплет нуклеотидов ДНК, кодирующий одну аминокислоту Б. Место присоединения аминокислоты к тРНК
В. Триплет нуклеотидов тРНК, комплементарный кодону мРНК
Г. Бессмысленный кодон мРНК Д. Триплет нуклеотидов РНК, кодирующий одну аминокислоту
51*. Выберите один неправильный ответ. Для образования Глу-тРНК глу необходимы:
В. тРНК глу Г. Глутамат
52*. Выберите один неправильный ответ. Энергия ГТФ при трансляции требуется для:
А. Образования пептидных связей
Б. Включения Мет-тРНК в пептидильный центр рибосомы В. Транслокации Г. Включения аминоацил-тРНК в центр связывания Д. Терминации
53*. Выберите один правильный ответ. На рибосоме адапторная РНК связана с:
Г. Аминоацил-тРНК-синтетазой Д. Факторами терминации
54*. Выберите один неправильный ответ.
При синтезе белка для образования инициирующего комплекса на рибосоме необходимы:
55*. Выберите один правильный ответ.
После включения в A-центр рибосомы кодона UAG наступает: А. Элонгация Б. Инициация
Г. Транслокация Д. Образование пептидной связи
56. Выполните «цепное» задание.
а) в синтезе белка у прокариотов и эукариотов участвуют:
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.



