Что это такое — теория симбиогенеза
Симбиогенез
Теория симбиогенеза (симбиотическая теория, эндосимбиотическая теория, теория эндосимбиоза) объясняет механизм возникновения некоторых органоидов эукариотической клетки — митохондрий, гидрогеносом и пластид.
Схема эволюции эукариотических клеток. 1 — образование двойной мембраны ядра, 2 — приобретение митохондрий, 3 — приобретение пластид, 4 — внедрение получившейся фотосинтезирующей эукариотической клетки в нефотосинтезирующую (например, в ходе эволюции криптофитовых водорослей), 5 — внедрение получившейся клетки снова в нефотосинтезирующую (например, при симбиозе этих водорослей с инфузориями). Цветом обозначен геном предков эукариот, митохондрий и пластид.
История cимбиогенеза
Теорию эндосимбиотического происхождения хлоропластов впервые предложил в 1883 году Андреас Шимпер, показавший их саморепликацию внутри клетки. Её возникновению предшествовал вывод А. С. Фаминцина и О. В. Баранецкого о двойственной природе лишайников — симбиотического комплекса гриба и водоросли (1867 год). К. С. Мережковский в 1905 году предложил само название «симбиогенез», впервые детально сформулировал теорию и даже создал на её основе новую систему органического мира. Фаминцин в 1907 году, опираясь на работы Шимпера, также пришёл к выводу, что хлоропласты являются симбионтами, как и водоросли в составе лишайников.
В 1920-е годы теория была развита Б. М. Козо-Полянским, было высказано предположение, что симбионтами являются и митохондрии. Затем долгое время о симбиогенезе практически не упоминали в научной литературе. Второе рождение расширенная и конкретизированная теория получила уже в работах Линн Маргулис начиная с 1960-х годов.
Симбиотическое происхождение митохондрий и пластид
В результате изучения последовательности оснований в митохондриальной ДНК были получены весьма убедительные доводы в пользу того, что митохондрии — это потомки аэробных бактерий (прокариот), родственных риккетсиям, поселившихся некогда в предковой эукариотической клетке и «научившимися» жить в ней в качестве симбионтов. Теперь митохондрии есть почти во всех эукариотических клетках, размножаться вне клетки они уже не способны.
Существуют свидетельства того, что первоначально эндосимбиотические предки митохондрий не могли ни импортировать белки, ни экспортировать АТФ. Вероятно, первоначально они получали от клетки-хозяина пируват, а выгода для хозяина состояла в обезвреживании аэробными симбионтами токсичного для нуклеоцитоплазмы кислорода.
Пластиды, подобно митохондриям, имеют свои собственные прокариотические ДНК и рибосомы. По-видимому, хлоропласты произошли от фотосинтезирующих бактерий, поселившихся в своё время в гетеротрофных клетках протистов, превратив их в автотрофные водоросли.
Доказательства симбиогенеза
Митохондрии и пластиды:
Проблемы симбиогенеза
Примеры эндосимбиозов
В наши дни существует ряд организмов, содержащих внутри своих клеток другие клетки в качестве эндосимбионтов. Они, однако, не являются сохранившимися до наших дней первичными эукариотами, у которых симбионты еще не интегрировались в единое целое и не потеряли своей индивидуальности. Тем не менее, они наглядно и убедительно показывают возможность симбиогенеза.
Гипотезы эндосимбиотического происхождения других органелл
Эндосимбиоз — наиболее широко признанная версия происхождения митохондрий и пластид. Но попытки объяснить подобным образом происхождение других органелл и структур клетки не находят достаточных доказательств и наталкиваются на обоснованную критику.
Клеточное ядро, нуклеоцитоплазма
Смешение у эукариот многих свойств, характерных для архей и бактерий, позволило предположить симбиотическое происхождение ядра от метаногенной архебактерии, внедрившейся в клетку миксобактерии. Гистоны, к примеру, обнаружены у эукариот и некоторых архей, кодирующие их гены весьма схожи. Другая гипотеза, объясняющая сочетание у эукариот молекулярных признаков архей и эубактерий, состоит в том, что на некотором этапе эволюции похожие на архей предки нуклеоцитоплазматического компонента эукариот приобрели способность к усиленному обмену генами с эубактериями путём горизонтального переноса генов.
В последнее десятилетие сформировалась также гипотеза вирусного эукариогенеза. В её основании лежит ряд сходств устройства генетического аппарата эукариот и вирусов: линейное строение ДНК, её тесное взаимодействие с белками и др. Было показано сходство ДНК-полимеразы эукариот и поксивирусов, что сделало именно их предков основными кандидатами на роль ядра.
Жгутики и реснички
Линн Маргулис в книге Symbiosis in Cell Evolution (1981) предположила в том числе происхождение жгутиков и ресничек от симбиотических спирохет. Несмотря на сходство размеров и строения указанных органелл и бактерий и существование Mixotricha paradoxa, использующей спирохет для движения, в жгутиках не было найдено никаких специфически спирохетных белков. Однако известен общий для всех бактерий и архей белок FtsZ, гомологичный тубулину и, возможно, являющийся его предшественником. Жгутики и реснички не обладают такими признаками бактериальных клеток, как замкнутая наружная мембрана, собственный белоксинтезирующий аппарат и способность к делению. Данные о наличии ДНК в базальных тельцах, появившиеся в 1990-е годы, были впоследствии опровергнуты. Увеличение числа базальных телец и гомологичных им центриолей происходит не путём деления, а путём достраивания нового органоида рядом со старым.
Пероксисомы
Кристиан де Дюв обнаружил пероксисомы в 1965 году. Ему же принадлежит предположение, что пероксисомы были первыми эндосимбионтами эукариотической клетки, позволившими ей выживать при нарастающем количестве свободного молекулярного кислорода в земной атмосфере. Пероксисомы, однако, в отличие от митохондрий и пластид, не имеют ни генетического материала, ни аппарата для синтеза белка. Было показано, что эти органеллы формируются в клетке de novo в ЭПР и нет никаких оснований считать их эндосимбионтами.
Биология. 11 класс
§ 13-1. Двумембранные органоиды
Двумембранными органоидами клеток являются митохондрии и пластиды.
Митохондрии — органоиды, в которых протекает кислородный этап клеточного дыхания (этот процесс будет подробно рассмотрен в следующей главе). В ходе кислородного этапа с участием О2 происходит расщепление и окисление органических соединений до неорганических веществ. При этом выделяется много энергии, которая используется для синтеза большого количества АТФ. Поэтому митохондрии иногда называют «энергетическими станциями» клетки.
Митохондрии являются динамичными органоидами. Они способны изменять свою форму, сливаться друг с другом, делиться, перемещаться в участки клетки с повышенным потреблением энергии. Митохондрии скапливаются преимущественно в тех частях клетки, где выше потребность в АТФ, например вблизи органоидов движения или миофибрилл.*
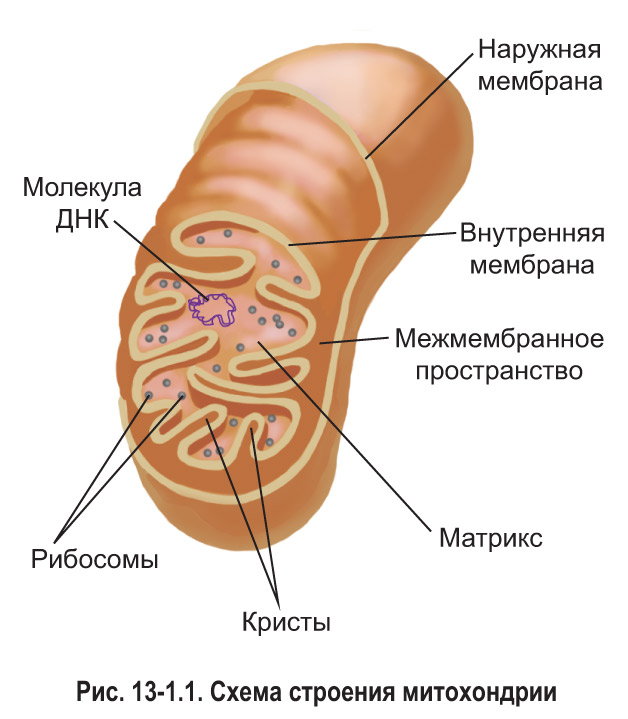 Каждая митохондрия ограничена двумя мембранами — наружной и внутренней, между которыми находится межмембранное пространство (рис. 13-1.1). Наружная мембрана митохондрии гладкая, не образует впячиваний и складок. Она отделяет органоид от гиалоплазмы и обладает высокой проницаемостью для ионов и небольших молекул. Внутренняя мембрана характеризуется гораздо меньшей проницаемостью. Она образует многочисленные складки — кристы, которые значительно увеличивают площадь ее поверхности. Внутренняя мембрана митохондрий содержит большое количество белков. В ее состав входят, например, ферменты, обеспечивающие синтез АТФ.
Каждая митохондрия ограничена двумя мембранами — наружной и внутренней, между которыми находится межмембранное пространство (рис. 13-1.1). Наружная мембрана митохондрии гладкая, не образует впячиваний и складок. Она отделяет органоид от гиалоплазмы и обладает высокой проницаемостью для ионов и небольших молекул. Внутренняя мембрана характеризуется гораздо меньшей проницаемостью. Она образует многочисленные складки — кристы, которые значительно увеличивают площадь ее поверхности. Внутренняя мембрана митохондрий содержит большое количество белков. В ее состав входят, например, ферменты, обеспечивающие синтез АТФ.
Содержимое митохондрии, ограниченное внутренней мембраной, называется матриксом. В матриксе содержатся различные неорганические и органические вещества, в том числе разнообразные ферменты, а также кольцевые молекулы ДНК и все виды РНК. Следовательно, митохондрии содержат собственную генетическую информацию. В их матриксе также находятся рибосомы, в которых осуществляется реализация этой информации, т. е. синтез белков. Митохондриальные *70S* рибосомы меньше по размерам, чем рибосомы, содержащиеся в гиалоплазме клетки. *ДНК митохондрии кодирует лишь небольшую часть белков, необходимых для функционирования этого органоида. Большинство митохондриальных белков кодируется ДНК, расположенной в ядре клетки. Такие белки синтезируются в 80S рибосомах в гиалоплазме, а затем транспортируются в митохондрию.*
Главная функция митохондрий — обеспечение клетки энергией в виде АТФ. *В клетке происходит постоянное обновление митохондрий. Новые митохондрии образуются в результате деления материнских. Этот процесс, как правило, протекает независимо от деления клетки и определяется ее энергетическими потребностями. Когда потребности клетки в энергии высоки, митохондрии интенсивно растут и размножаются путем деления. Если потребление энергии низкое, часть митохондрий может разрушаться или переходить в неактивное состояние.*
Биология клетки/Часть 1. Клетка как она есть/1/2
Last Universal Common Ancestor (LUCA; последний универсальный общий предок, англ.)
Содержание
Происхождение клеток и происхождение эукариот [ править ]
Клетки эукариот — сложные генетические системы, имеющие другой уровень организации, чем прокариотические клетки.
Во-первых, у эукариот намного сложнее устроена система регуляции работы генов. Это позволяет им приспосабливаться к более разнообразным условиям среды без изменений в их ДНК. Многоклеточным организмам это позволяет создавать разные типы клеток, имеющие один и тот же генотип.
Во-вторых, у подавляющего большинства эукариот есть митохондрии, а у многих ещё и пластиды. Согласно общепризнанной в настоящее время точке зрения, эти органоиды имеют симбиотическое происхождения. Их предками были бактерии, которые вступили в симбиоз с предками эукариот.
Теория симбиотического (симбиогенного) происхождения органоидов клетки получила название теории симбиогенеза.
Теория симбиогенеза [ править ]
Теорию симбиогенеза создали русские учёные, а ввела в современный научный обиход американский исследователь Линн Маргелис [ править ]
Теория эндосимбиотического происхождения хлоропластов впервые была предложена в 1883 году Андреасом Шимпером, показавшим их саморепликацию внутри клетки. Её возникновению предшествовал вывод А. С. Фаминцина и О. В. Баранецкого о двойственой природе лишайников — симбиотического комплекса гриба и водоросли (1867 год). К. С. Мережковский в 1905 году предложил само название «симбиогенез», впервые детально сформулировал теорию и даже создал на её основе новую систему органического мира.
Фаминцин в 1907 году, опираясь на работы Шимпера, также пришел к выводу, что хлоропласты являются симбионтами, как и в случае с водорослями лишайника.
В 1920-е годы теорию симбиогенеза развивал российский ботаник Б.М. Козо-Полянский [1]. Его работы, публиковавшиеся на русском языке, остались полностью неизвестными на западе. Еще более четко, чем Мережковский, Козо-Полянский осознал и обозначил границу между про- и эукариотами. Теорию симбиогенеза он распространил на митохондрии (и другие органоиды, включая ядро). Он также отчасти предвосхитил современные представления о внеядерной наследственности. Хотя у симбиогенеза находились адепты и в 30-40-е годы, в целом эту теорию большинство ученых считали ненаучной фантастикой. Второе рождение расширенная и конкретизированная теория получила в работах Линн Маргулис (Маргелис) начиная с 1960-х гг. Маргелис высказала предположение о симбиотическом происхождении не только митохондрий и хлоропластов, но и жгутиков (от спирохет), которое в дальнейшем не получило подтверждения.
Согласно теории симбиогенеза, митохондрии и хлоропласты — потомки бактерий, вступившие в симбиоз с предками эукариот [ править ]
Современная теория симбиогенеза утверждает, что митохондрии и хлоропласты — потомки определённых групп бактерий, которые вступили в симбиоз с предками современных эукариот. В ходе эволюции бактерии-эндосимбионты превратились в полуавтономные органоиды. Они сохранили способность синтезировать некоторые белки автономно от клетки-хозяина и способность размножаться путем деления. Но значительная часть генетического материала митохондрий и хлоропластов переместилась в ядро. В результате эти органоиды утратили способность размножаться вне клетки-хозяина, свойственную многим симбиотическим бактериям.
Митохондрии и хлоропласты имеют признаки минимальной клетки — замкнутую наружную мембрану, ДНК, белоксинтезирующий аппарат и размножение делением [ править ]
Анализ нуклеотидных последовательностей ДНК митохондрий и хлоропластов свидетельствует о том, что каждый из этих органоидов был приобретены предками эукариот однократно. Все митохондрии всех современных эукариот имеют одного общего предка, сходного с современными альфа-протеобактериями. Все хлоропласты тоже имеют одного общего предка, сходного с современными цианобактериями. Особенно близки к предкам хлоропластов по строению мембран и составу фотосинтетических пигментов бактерии рода прохлорон (Prochloron).
Отдельного комментария заслуживает наличие двух мембран у митохондрии и «типичных» хлоропластов. Широко распространенное заблуждение – бактериальное происхождение одной внутренней мембраны и возникновение внешней мембраны из мембраны пищеварительной вакуоли. Оно настолько укоренилось, что тиражируется во вполне солидных и уважаемых источниках (см., например, h учебник Альбертса «Молекулярная биология клетки», 5-е издание).
На самом же деле наружная мембрана митохондрий и хлоропластов гомологична наружной мембране их предков – грамотрицательных бактерий. «Бегство» из пищеварительной вакуоли – обычная тактика многих внутриклеточных бактерий. Так ведут себя многие возбудители болезней человека – листерии, риккетсии и др.
Во-первых, если бы наружная мембрана была гомологична мембране фагосомы, то белки в нее доставлялись путем везикулярного транспорта. Если это и происходит, то в виде исключения; в целом митохондрии и пластиды изолированы от «единой» мембранной системы, белки и липиды доставляются в них с помощью особых механизмов. Во-вторых, только в наружной мембране митохондрий, хлоропластов и грамм-отрицательных бактерий встречается уникальное семейство белков из группы бета-бочек, и только в этих мембранах работает уникальный, имеющий общее происхождение механизм встраивания этих белков.
Геном полуавтономных органелл резко уменьшен, многие их гены перенесены в ядро [ править ]
Наиболее полный из изученных геномов митохондрий у протиста Reclinomonas. В ДНК его митохондрии содержится более 60 «белковых» генов. У этого протиста в митохондриальной ДНК закодированы 27 рибосомальных белков, а также 23 белка, участвующих в работе электронтранспортной цепи и в окислительном фосфорилировании. Кроме того, у этого же протиста в мтДНК закодированы как минимум пять белков, участвующих в транскрипции (субъединицы РНК-полимеразы) и трансляции.
У человека (как и у большинства животных) в ДНК митохондрий кодируются рРНК митохондриальных рибосом и большинство типов митохондриальных тРНК. При этом геном митохондрий человека и других млекопитающих содержит всего 13 «белковых» генов из примерно 1500 генов, кодирующих белки митохондриального протеома.
Перенос генов из органоидов в ядро
Геном митохондрий имеет необычные черты, в том числе отклонения от универсального генетического кода [ править ]
Необычные черты генома митохондрий
В иРНК митохондрий особенно часто встречаются вариации стандартного генетического кода, которые приведены в таблице.
| Пример | Кодон | Обычное значение | Читается как: |
|---|---|---|---|
| Митохондрии, в частности у Saccharomyces cerevisiae | CU(U, C, A, G) | Лейцин | Серин |
| Митохондрии высших растений | CGG | Аргинин | Триптофан |
| Митохондрии (у всех без исключения исследованных организмов) | UGA | Стоп | Триптофан |
| Митохондирии млекопитающих, дрозофилы, S. cerevisiae и многих простейших | AUA | Изолейцин | Метионин = Старт |
| Митохондрии млекопитающих | AGC, AGU | Серин | Стоп |
| Митохондрии дрозофилы | AGA | Аргинин | Стоп |
| Митохондрии млекопитающих | AG(A, G) | Аргинин | Стоп |
У большинства изученных организмов митохондрии содержат только кольцевые молекулы ДНК, у некоторых растений одновременно присутствуют и кольцевые, и линейные молекулы, а у ряда протистов (например, инфузорий) имеются только линейные молекулы.
Митохондрии млекопитающих обычно содержат от двух до десяти идентичных копий кольцевых молекул ДНК.
У растений каждая митохондрия содержит несколько молекул ДНК разного размера, которые способны к рекомбинации.
Митохондрии кинетопластид имеют необычное строение и генетический аппарат [ править ]
У протистов из отряда кинетопластид (например, у трипаносом) в особом участке митохондрии (кинетопласте) содержится два типа молекул ДНК — идентичные макси-кольца (20-50 штук) длиной около 21 т.п.о. и мини-кольца (20 000 — 55 000 штук, около 300 разновидностей, средняя длина около 1000 п.о.). Все кольца соединены в единую сеть (катенаны), которая разрушается и восстанавливается при каждом цикле репликации. Макси-кольца гомологичны митохондриальной ДНК других организмов. Каждое мини-кольцо содержит четыре сходных консервативных участка и четыре уникальных гипервариабельных участка.
В мини-кольцах закодированы короткие молекулы направляющих РНК (guideRNA), которые осуществляют редактирование РНК, транскрибируемых с генов макси-колец.
Пластиды многих водорослей отличаются от пластид высших растений строением и составом пигментов [ править ]
В большинстве учебников ботаники и физиологии растений описаны пластиды (хлоропласты), характерные для цветковых растений. Они имеют две наружные мембраны, тилакоиды, собранные в стопки — граны, и два основных фотосинтетических пигмента — хлорофилл а и хлорофилл b.Но на самом деле пластиды гораздо разнообразнее и по строению, и по составу пигментов.
Первичные пластиды характерны, кроме высших растений, для зеленых и харовых водорослей, красных водорослей и глаукофитовых. Все эти пластиды окружены двумя мембранами. Считается. что они были приобретены в результате симбиоза гетеротрофных жгутиковых эукариот с предками хлоропластов — цианобактериями.
При этом и у этих групп водорослей пластиды заметно различаются. У зеленых и харовых водорослей пластиды по строению и составу пигментов такие же, как у высших растений. Считается, что высшие растения произошли от водорослей, сходных с харовыми. По строению их хлоропласты наиболее сходны с цианобактерией Prochloron. Эти пластиды содержат две формы хлорофилла — хлорофилл а и хлорофилл b. Те же формы хлорофилла имеет и Prochloron.
У красных и глаукофитовых водорослей тилакоиды не собраны в граны, а на их поверхности сидят особые тельца — фикобилисомы. Они состоят из белков фикобилипротеинов. Окраску им придают входящие в их состав хромофоры фикобилины: фикоэритрин — красную, а фикоцианин — голубоватую. Они улавливают свет и передают его хлорофиллу а. У красных водорослей и некоторых цианобактерий есть также хлорофилл d.
По строению и составу пигментов эти хлоропласты наиболее сходны с большинством видов цианобактерий. При этом хлоропласты глаукофитовых водорослей имеют удивительную особенность: между двумя мембранами у них есть слой пептидогликана — вещества, образующего клеточные стенки бактерий. Эти хлоропласты, называемые часто цианеллы, рассматривают как «живые ископаемые» — эндосимбиотические органеллы, сохранившие примитивные предковые признаки.
Вторичные пластиды характерны для большинства групп водорослей. Они были приобретены в результате эндосимбиоза гетеротрофных протистов с автотрофными протистами, то есть в результате поглощения хищником-эукариотом другого эукариота — автотрофа, уже имевшего первичные пластиды. Исходно вторичные пластиды окружены четырьмя мембранами (снаружи внутрь — мембрана пищеварительной вакуоли, плазмалемма эукариотической клетки, две мембраны первичной пластиды). Впоследствии у многих групп водорослей одна или две их четырех мембран вторичных пластид редуцировались, а у некоторых все четыре мембраны сохранились.
Наиболее близки к исходному строению хлоропласты криптофитовых водорослей и хлорарахиевых водорослей. У этих небольших групп одноклеточных водорослей хлоропласты окружены четырьмя мембранами. У криптофитовых они содержат хлорофилл а, хлорофилл с и фикобилипротеины. У хлорарахниевых присутствуют хлорофилл а и хлорофилл b. Между второй и третьей мембраной находится нуклеоморф — рудиментарное ядро с сильно уменьшенным ядерным геномом.
Сейчас секвенированы полные геномы всех компартментов обоих групп водорослей. Оба исследованных нуклеоморфа содержат по три маленькие хромосомы (у изученной криптофитовой водоросли общий размер генома всего 550.000 п.н.). Из белковых генов нуклеоморфов всего примерно по 30 генов кодируют белки для первичной пластиды; остальные — это в основном «гены домашнего хозяйства» Их тоже не хватает: в перипластидное пространство (то есть в эукариота-симбионта) попадает из цитоплазмы хозяина более 2000 белков у криптофитовых и около 1000 у хлорарахниевых. В свою очередь, из нуклеоморфов в ядро переместились сотни генов. Геном нуклеоморфа у криптофитовых очень компактный: в нем короткие интроны, почти нет спейсеров, а 91 % последовательностей — кодирующие, как у прокариот!
У гаптофитовых и диатомовых водорослей, недавно открытого фототрофного протиста Chromera velia и у ее ближайших родственников споровиков (Apycomplexa) есть четырехмембранные пластиды, утратившие нуклеоморф. У других водорослей (например, динофлагеллят и эвгленовых) встречаются трехмембранные хлоропласты со своеобразным набором хлорофиллов. Наличие трех мембран можно объяснить тем, что после двойного симбиоза (как у криптофитовых) одна из мембран редуцировалась.
По-видимому, в данном случае это была наружная мембрана клетки эукариотического симбионта; сохранились обе мембраны первичной пластиды и мембрана фагосомы.
Кем же были предки вторичных пластид? Сейчас считается, что хлорарахниевые и эвгленовые независимо друг от друга поработили зеленые водоросли. Все остальные группы, имеющие вторичные пластиды, приобрели в свое время в качестве симбионта красную водоросль
[5]Patrick J. Keeling. Diversity and evolutionary history of plastids and their hosts.\
Симбиогенез


Теория симбиогене́за (симбиотическая теория, эндосимбиотическая теория, теория эндосимбиоза) объясняет механизм возникновения некоторых органоидов эукариотической клетки — митохондрий, гидрогеносом и пластид.
Содержание
История
В 1920-е годы теория была развита Б. М. Козо-Полянским, было высказано предположение, что симбионтами являются и митохондрии. Затем долгое время о симбиогенезе практически не упоминали в научной литературе. Второе рождение расширенная и конкретизированная теория получила уже в работах Линн Маргулис начиная с 1960-х годов.
Симбиотическое происхождение митохондрий и пластид
В результате изучения последовательности оснований в митохондриальной ДНК были получены весьма убедительные доводы в пользу того, что митохондрии — это потомки аэробных бактерий (прокариот), родственных риккетсиям, поселившихся некогда в предковой эукариотической клетке и «научившимися» жить в ней в качестве симбионтов. Теперь митохондрии есть почти во всех эукариотических клетках, размножаться вне клетки они уже не способны.
Пластиды, подобно митохондриям, имеют свои собственные прокариотические ДНК и рибосомы. По-видимому, хлоропласты произошли от фотосинтезирующих бактерий, поселившихся в свое время в гетеротрофных клетках протистов, превратив их в автотрофные водоросли.
Доказательства
Проблемы
Примеры эндосимбиозов
В наши дни существует ряд организмов, содержащих внутри своих клеток другие клетки в качестве эндосимбионтов. Они, однако, не являются сохранившимися до наших дней первичными эукариотами, у которых симбионты еще не интегрировались в единое целое и не потеряли своей индивидуальности. Тем не менее, они наглядно и убедительно показывают возможность симбиогенеза.
Гипотезы эндосимбиотического происхождения других органелл
Эндосимбиоз — наиболее широко признанная версия происхождения митохондрий и пластид. Но попытки объяснить подобным образом происхождение других органелл и структур клетки не находят достаточных доказательств и наталкиваются на обоснованную критику.
Клеточное ядро, нуклеоцитоплазма
Смешение у эукариот многих свойств, характерных для архей и бактерий, позволило предпололожить симбиотическое происхождение ядра от метаногенной архебактерии, внедрившейся в клетку миксобактерии. Гистоны, к примеру, обнаружены у эукариот и некоторых архей, кодирующие их гены весьма схожи. Другая гипотеза, объясняющая сочетание у эукариот молекулярных признаков архей и эубактерий, состоит в том, что на некотором этапе эволюции похожие на архей предки нуклеоцитоплазматического компонента эукариот приобрели способность к усиленному обмену генами с эубактериями путём горизонтального переноса генов [5]
Жгутики и реснички
Линн Маргулис в книге Symbiosis in Cell Evolution (1981) предположила в том числе происхождение жгутиков и ресничек от симбиотических спирохет. Несмотря на сходство размеров и строения указанных органелл и бактерий и существование Mixotricha paradoxa, использующей спирохет для движения, в жгутиках не было найдено никаких специфически спирохетных белков. Однако известен общий для всех бактерий и архей белок FtsZ, гомологичный тубулину и, возможно, являющийся его предшественником. Жгутики и реснички не обладают такими признаками бактериальных клеток, как замкнутая наружная мембрана, собственный белоксинтезирующий аппарат и способность к делению. Данные о наличии ДНК в базальных тельцах, появившиеся в 1990-е годы, были впоследствии опровергнуты. Увеличение числа базальных телец и гомологичных им центриолей происходит не путем деления, а путем достраивания нового органоида рядом со старым.



