Кроссинговер
Кроссинговер

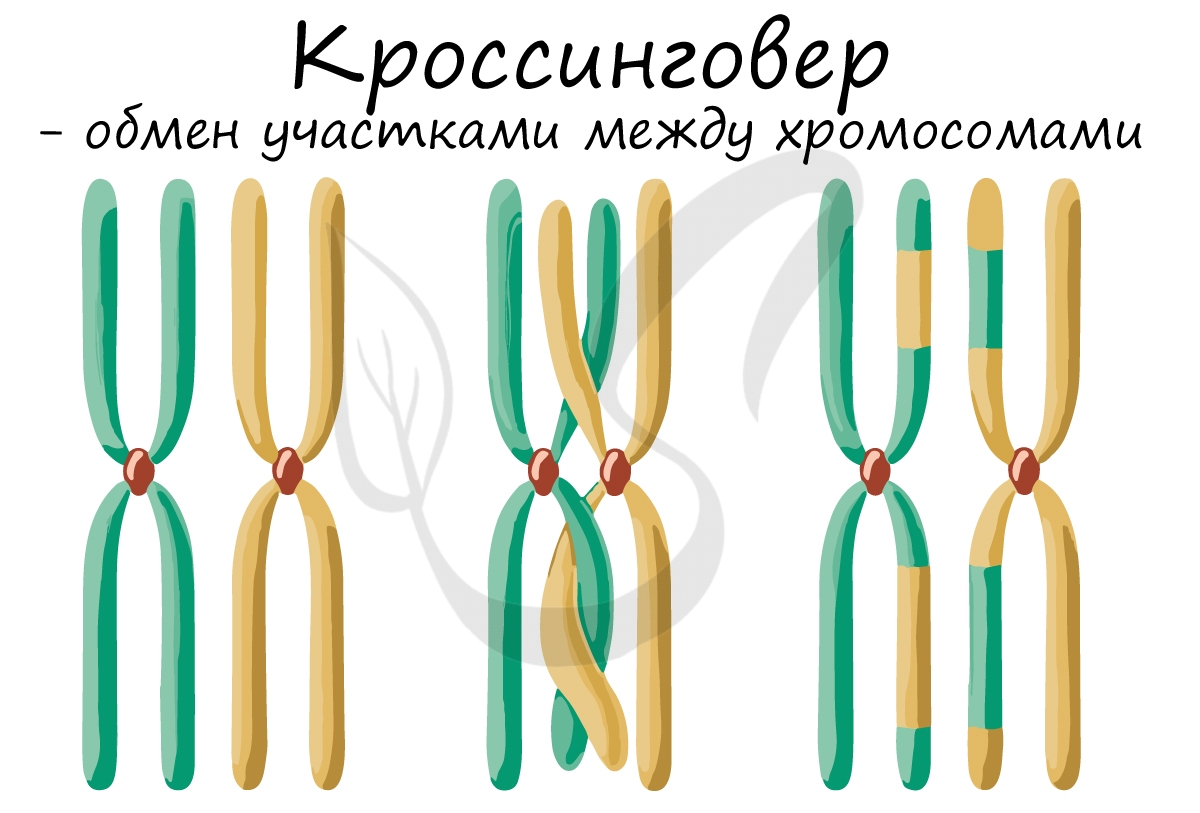
Кроссинговер — перекрест, взаимный обмен гомологичными участками гомологичных хромосом в результате разрыва и соединения в новом порядке их нитей — хроматид; приводит к новым комбинациям аллелей разных генов. Важнейший механизм, обеспечивающий комбинативную изменчивость в популяциях и тем самым дающий материал для естественного отбора. Протекает в мейотически, реже — в митотически делящихся клетках. Может приводить к перекомбинации больших|больших участков хромосомы с несколькими генами или частей одного гена (внутригенный кроссинговер), обеих нитей молекулы ДНК или только одной. Частота кроссинговера между генами отражает расстояние между ними в хромосоме. Иными слова|словами, в паре гомологичных хромосом между несестринскими хроматидами происходит обмен гомологичными участками. Поскольку в паре хромосом одна хромосома происходит от матери, а другая — от отца, процесс кроссинговера ведёт к внутрихромосомным рекомбинациям наследственности. Молекулярный механизм кроссинговера окончательно не выяснен.
Кроссинговер

Кроссинговер (crossing-over): обмен генетического материала междухромосомами, как результат «разрыва» и соединения хромосом;процесс обмена участками хромосом при перекресте хромосом (рис. 118, Б4).
Посколькукроссинговер — взаимный обмен гомологичными участками хромосом междугомологичными (парными|парными) хромосомами исходных гаплоидных наборов- особи имеют новые, различающиеся между собой генотипы. При этомдостигается перекомбинация наследственных свойств родителей, чтоувеличивает изменчивость и даёт более богатый материал для естественногоотбора.
Гены перемешиваются благодаря слиянию гамет двух различныхособей|особей, однако генетические изменения осуществляются не только этим путём.Никакие два потомка одних и тех же родителей (если только это не идентичныеблизнецы) не будут абсолютно одинаковыми. Во время мейоза осуществляются два различных видапересортировки генов.
На этой стадии мейозагомологи в каждой паре (или бивалент) остаются связанными друг с другом поменьшей мере одной хиазмой. Во многих бивалентах бывает большее числохиазм, так как возможны множественные перекресты между гомологами
Видео по теме : Кроссинговер
Кроссинговер

23.3.1. Кроссинговер и частота рекомбинаций
В 1909 г. бельгийский цитолог Янссенс наблюдал образование хиазм во время профазы I мейоза (см. разд. 22.3). Генетическое значение этого процесса разъяснил Морган, высказавший мнение, что кроссинговер (обмен аллелями) происходит в результате разрыва и рекомбинации гомологичных хромосом во время образования хиазм. В дальнейшем сопоставление цитологических данных с данными о соотношениях рекомбинантных фенотипов подтвердило, что обмен генетическим материалом в мейозе происходит почти буквально между всеми гомологичными хромосомами. Аллели, входящие в группы сцепления у родительских особей|особей, разделяются и образуют новые сочетания, которые попадают|попадают в гаметы,- процесс, называемый генетической рекомбинацией. Потомков, которые получаются из таких гамет с «новыми» сочетаниями аллелей, называют рекомбинантами. Таким образом, кроссинговер представляет собой важный источник генетической изменчивости, наблюдаемой в популяциях.
Для иллюстрации принципа кроссинговера можно рассмотреть поведение пары|пары гомологичных хромосом дрозофилы, несущих аллели серой окраски тела|тела и длинных крыльев (оба аллеля доминантные) и чёрной окраски тела|тела и зачаточных крыльев (оба аллеля рецессивные), во время образования хиазм. Скрещивание между гомозиготным серым длиннокрылым самцом и гомозиготной чёрной самкой с зачаточными крыльями дало в F1 гетерозиготных потомков с серым телом и длинными крыльями (рис. 23.10).
Рис. 23.10. Генетическое объяснение процесса кроссинговера и появления рекомбинантных генотипов. Подсчитав число особей|особей, у которых выявляется рекомбинация (х), и общее|общее число особей|особей (у), можно вычислить частоту рекомбинаций по формуле: Частота рекомбинации (%) =
При возвратном скрещивании мух из поколения F1 с гомозиготными двойными рецессивами были получены следующие результаты:
Как показывают эти результаты, гены, определяющие окраску тела|тела и длину крыльев, сцеплены. (Вспомните, что если бы эти гены находились в разных хромосомах и поэтому распределялись случайным образом, то при дигибридном скрещивании гетерозиготы F1 с гомозиготой по двум рецессив-ным признакам получилось бы соотношение фенотипов 1:1:1:1.) Из приведённых цифр можно вычислить частоту рекомбинации генов, определяющих окраску тела|тела и длину крыльев.
Частота рекомбинаций вычисляется по формуле
В нашем примере частота рекомбинации равна
Это значение соответствует числу рекомбинаций, происходящих при образовании гамет. Один из учеников Моргана, А. X. Стертевант, высказал мысль, что частоты|частоты рекомбинаций свидетельствуют о линейном расположении генов вдоль хромосомы. Ещё более важное предположение Стертеванта состояло в том, что частота рекомбинаций отражает относительное расположение генов в хромосоме: чем дальше друг от друга находятся сцепленные гены, тем больше вероятность того, что между ними произойдёт кроссинговер, т.е. тем выше частота рекомбинантов (рис. 23.11).
Рис. 23.11. А В и С — три генных локуса, расположенных в одной хромосоме. Вероятность кроссинговера и разделения для генов А и С выше, чем для генов В и С или А и В, так как частота кроссинговера зависит от расстояния между генами
23.8. На приведённой ниже схеме изображены локусы двенадцати аллелей, расположенные в паре хромосом. Показаны их относительные расстояния от центромеры.
а) Как называют изображённые здесь хромосомы?
б) Между какими двумя локусами чаще всего будет происходить кроссинговер?
в) Произойдёт ли кроссинговер между генами, определяющими цвет глаз и форму антенн? Обоснуйте свой ответ.
Митоз и мейоз
Жизненный цикл клетки (клеточный цикл)
С момента появления клетки и до ее смерти в результате апоптоза (программируемой клеточной гибели) непрерывно продолжается жизненный цикл клетки.
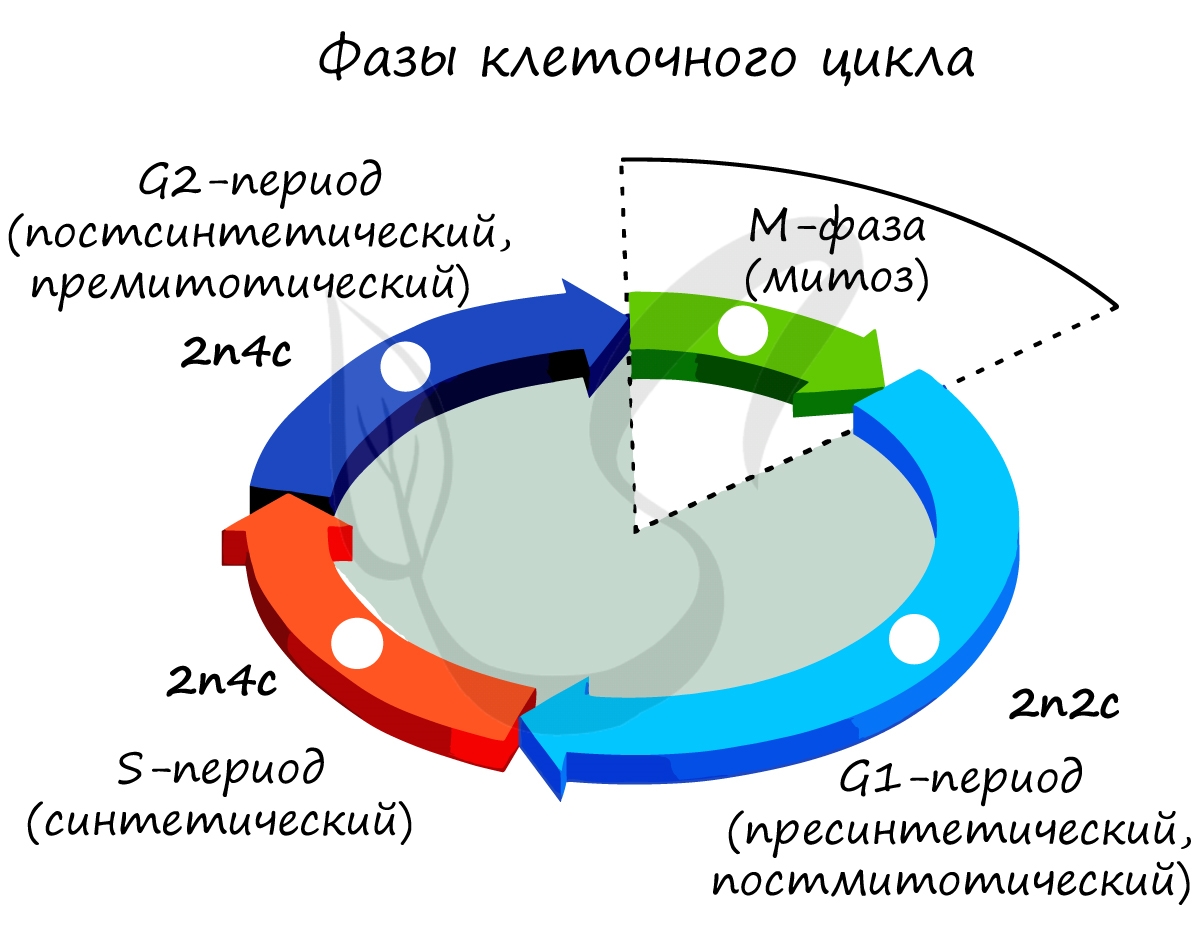
Интенсивно образуются рибосомы, синтезируется АТФ и все виды РНК, ферменты, клетка растет.
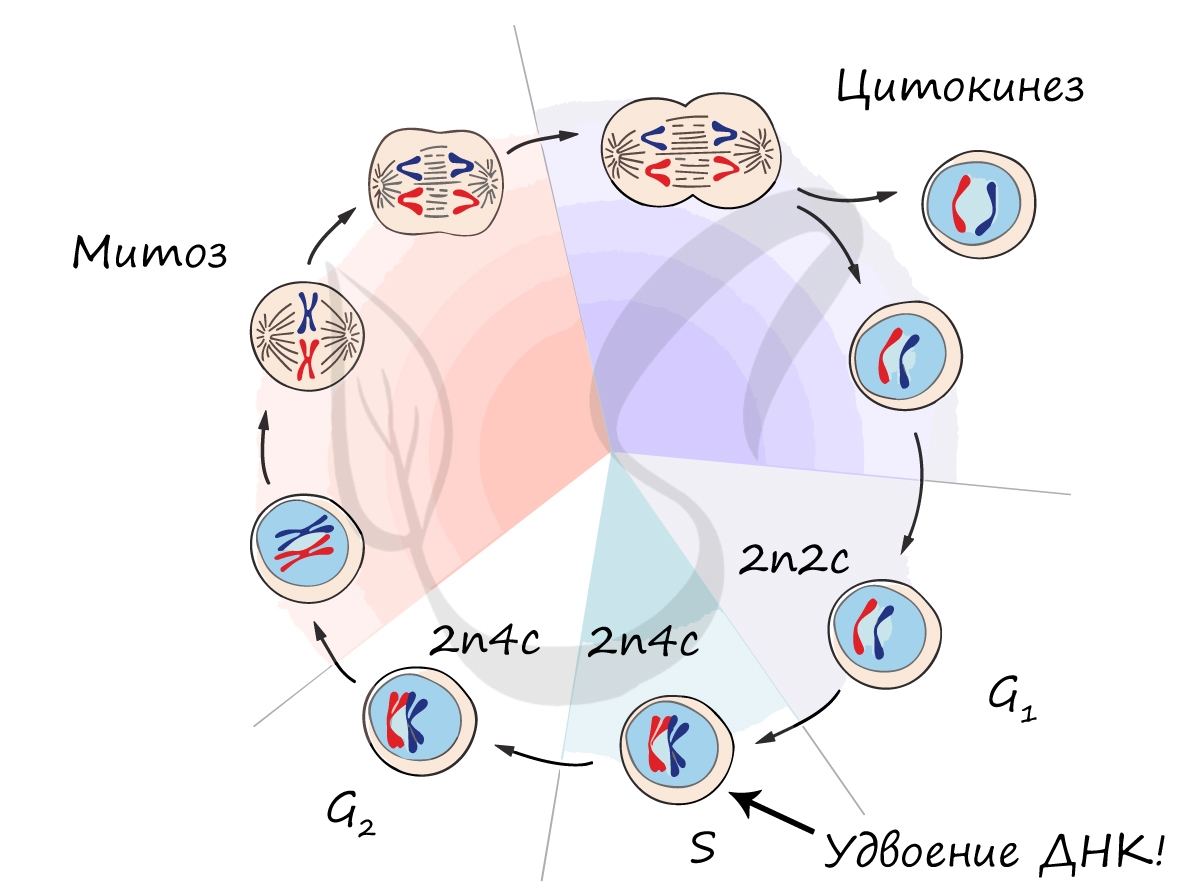
Митоз является непрямым способом деления клетки, наиболее распространенным среди эукариотических организмов. По продолжительности занимает около 1 часа. К митозу клетка готовится в период интерфазы путем синтеза белков, АТФ и удвоения молекулы ДНК в синтетическом периоде.
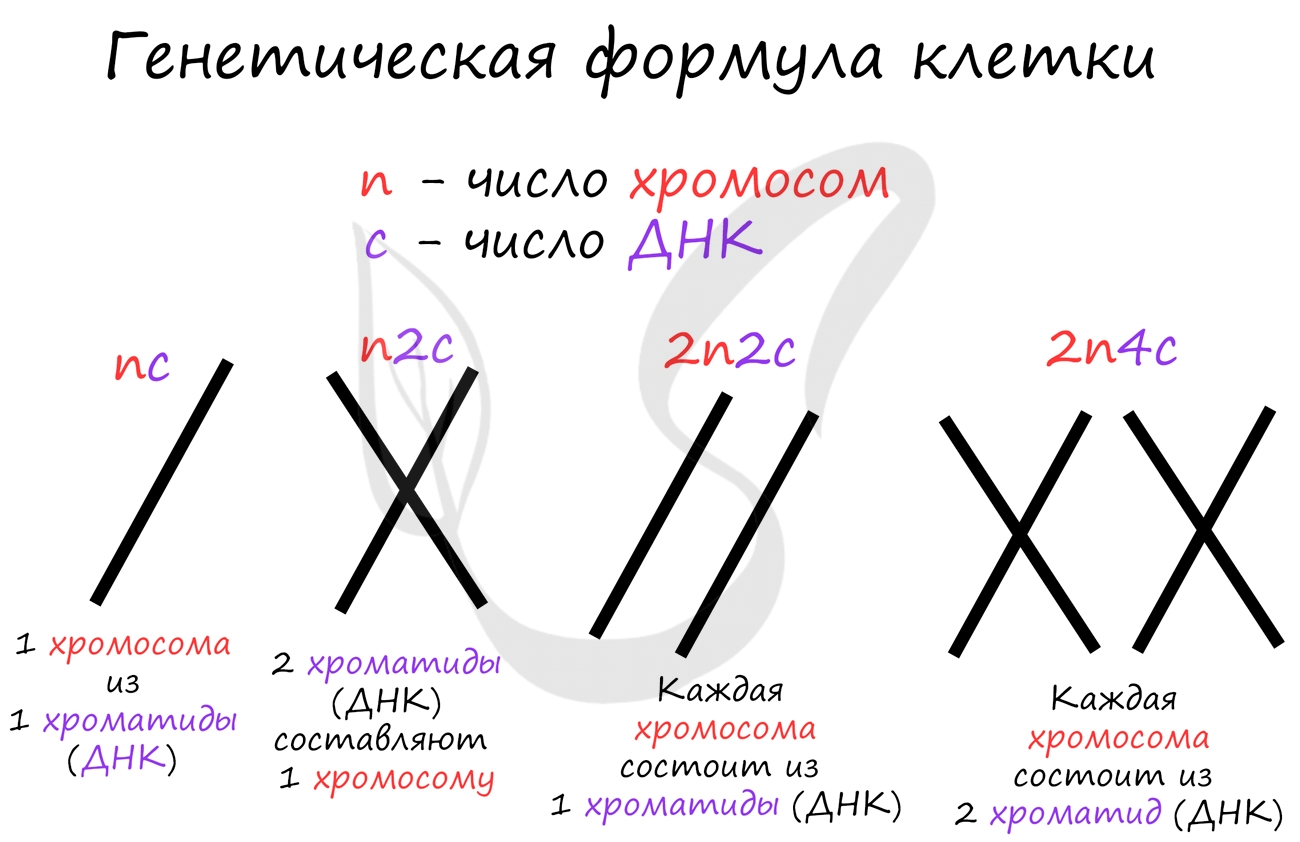
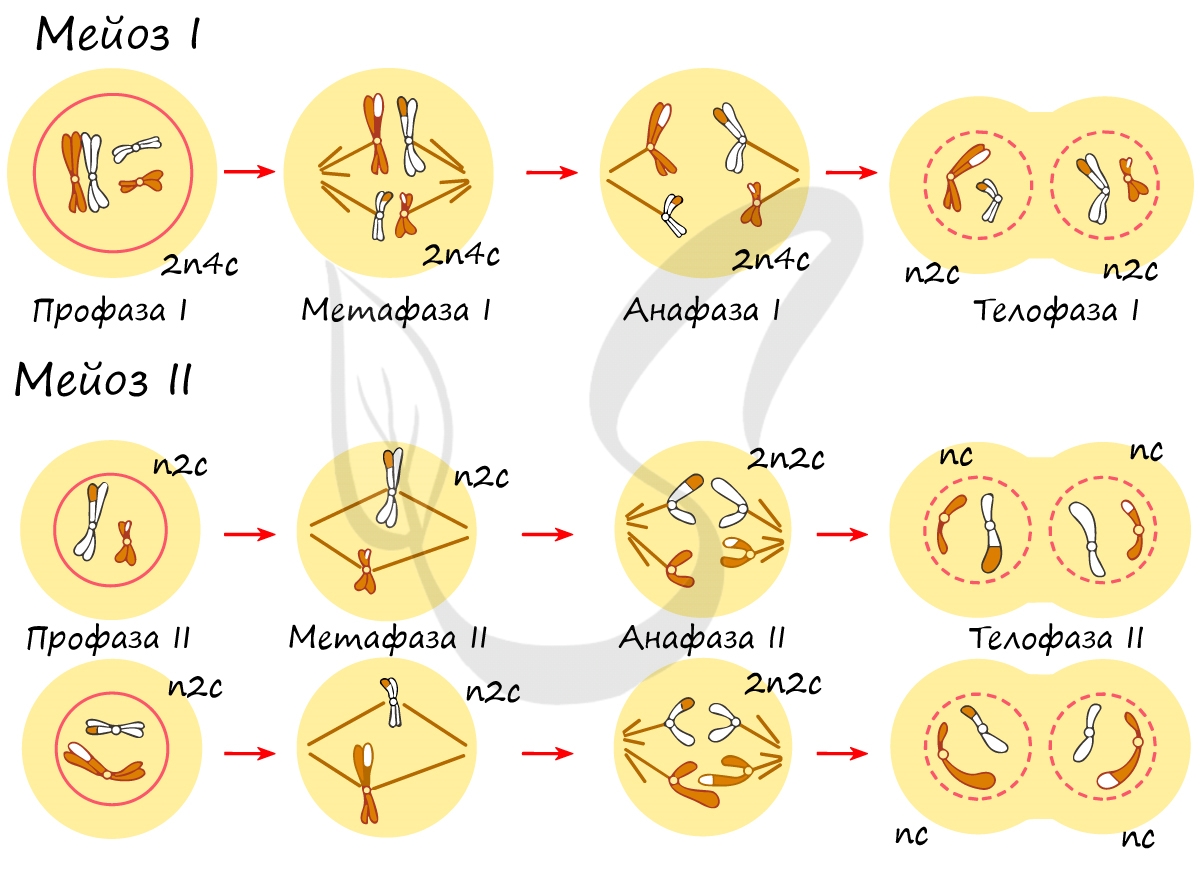
Митоз состоит из 4 фаз, которые мы далее детально рассмотрим: профаза, метафаза, анафаза, телофаза. Напомню, что клетка вступает в митоз с уже удвоенным (в синтетическом периоде) количеством ДНК. Мы рассмотрим митоз на примере клетки с набором хромосом и ДНК 2n4c.
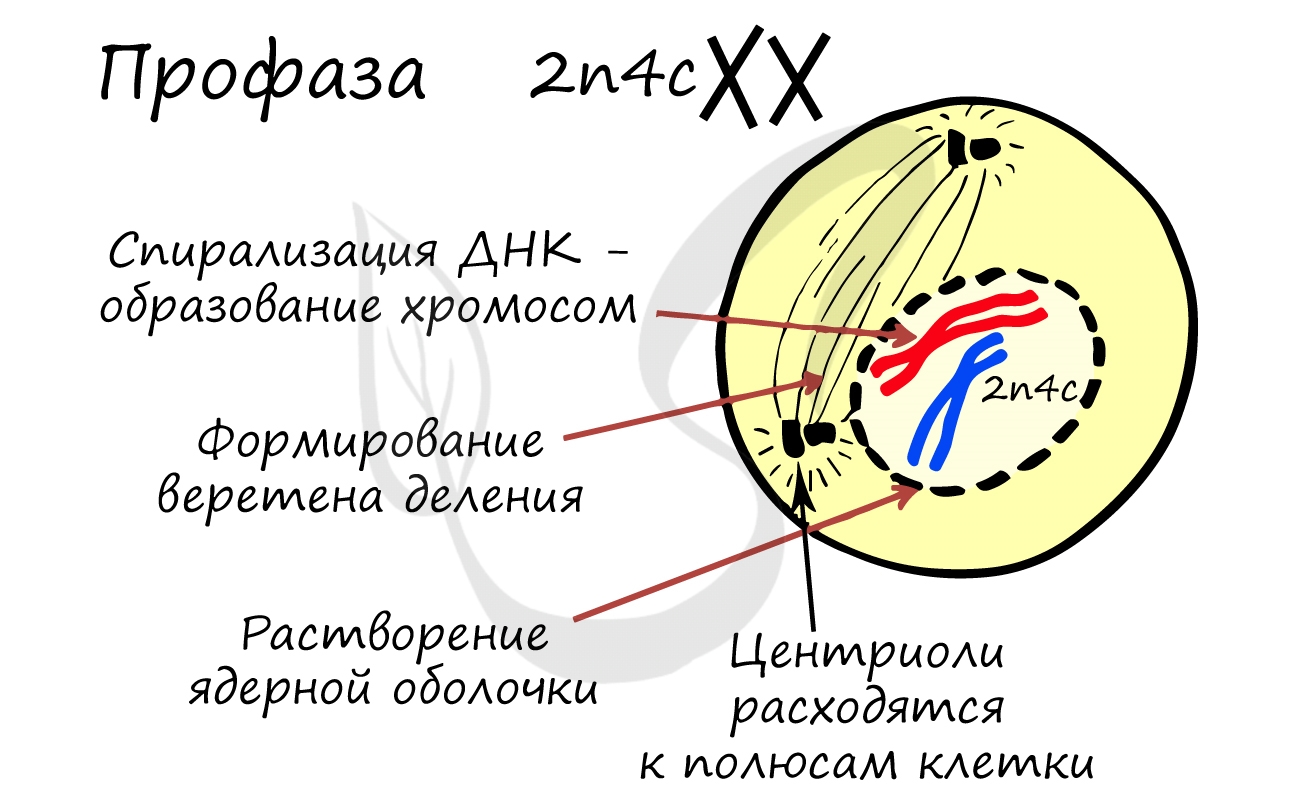
ДНК максимально спирализована в хромосомы, которые располагаются на экваторе клетки. Каждая хромосома состоит из двух хроматид, соединенных центромерой (кинетохором). Нити веретена деления прикрепляются к центромерам хромосом (если точнее, прикрепляются к кинетохору центромеры).
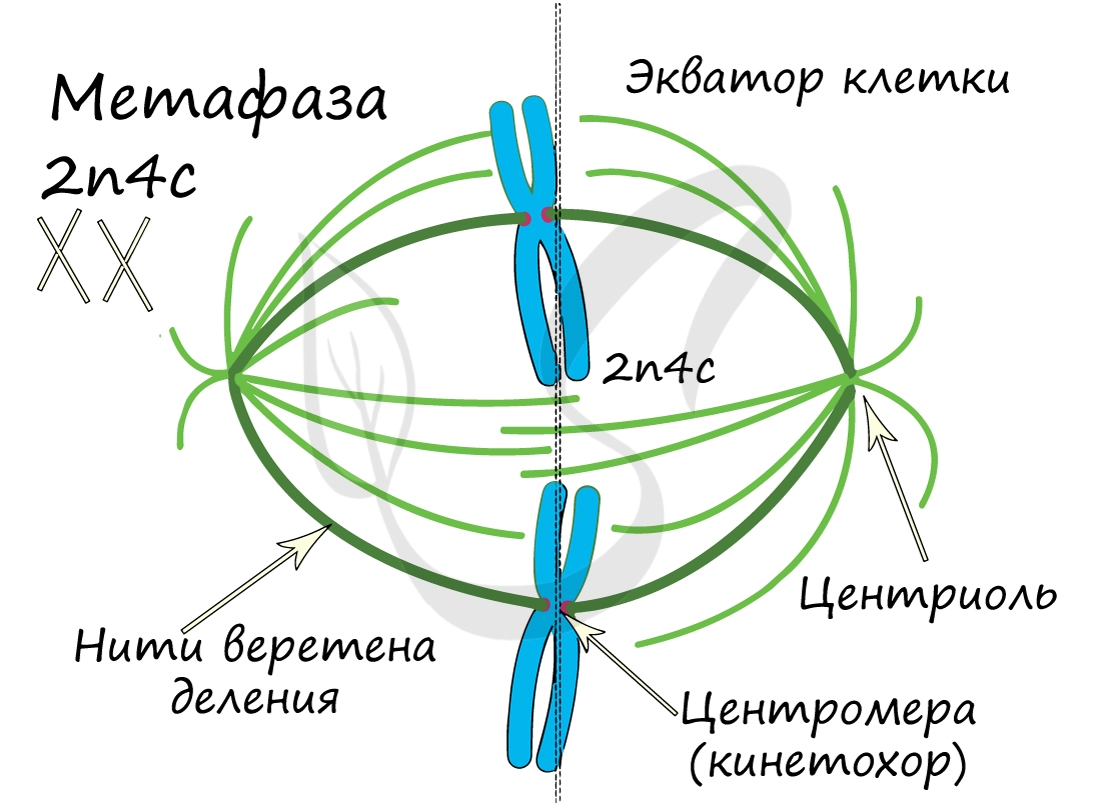
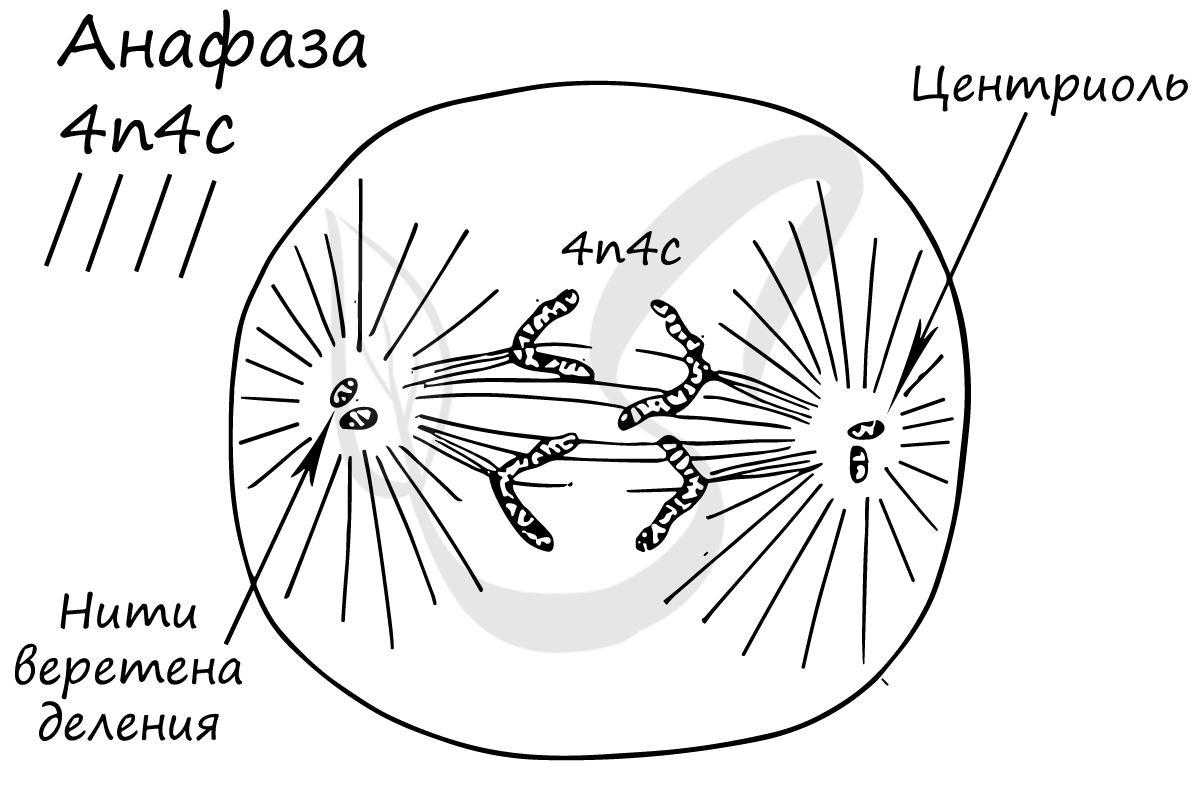
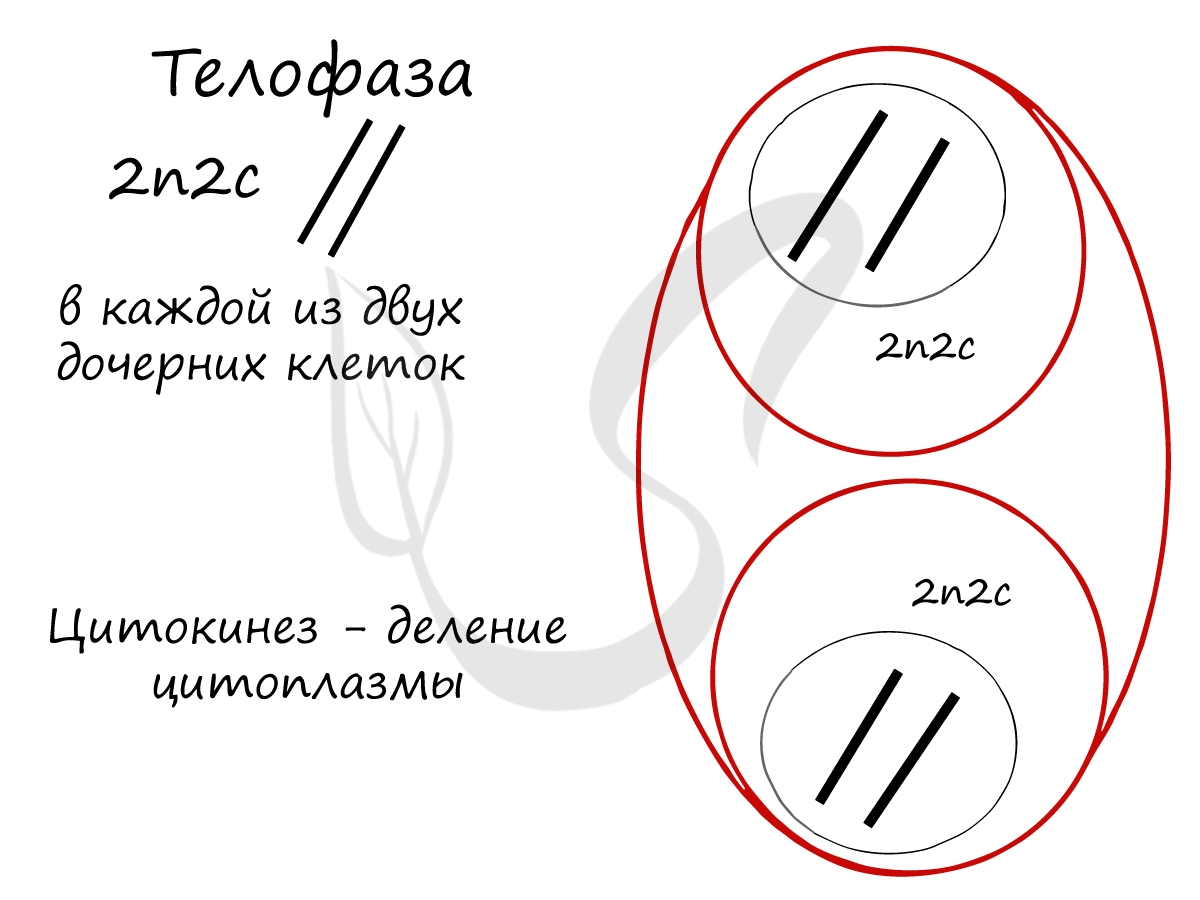
Попробуйте самостоятельно вспомнить фазы митоза и описать события, которые в них происходят. Особенное внимание уделите состоянию хромосом, подчеркните сколько в них содержится молекул ДНК (хроматид).
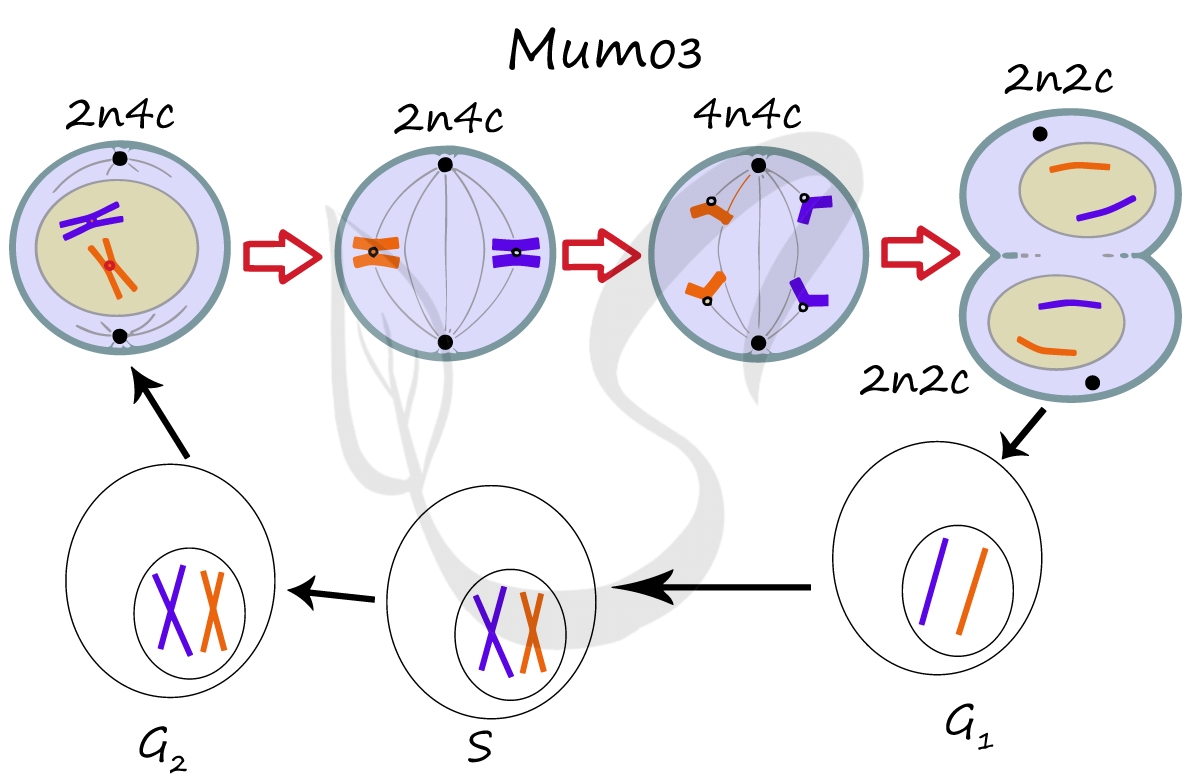
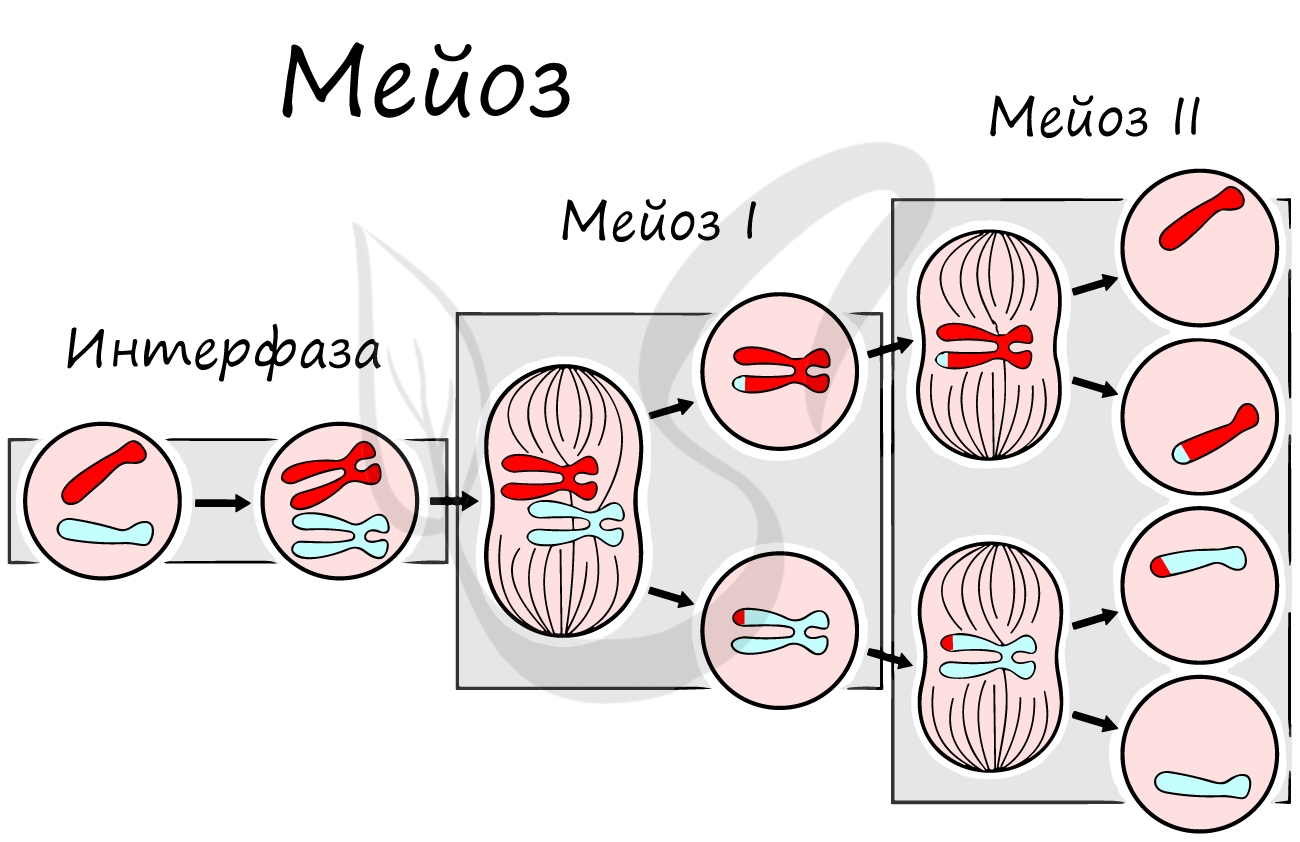
Мейоз
В результате мейоза из диплоидных клеток (2n) получаются гаплоидные (n). Мейоз состоит из двух последовательных делений, между которыми практически отсутствует пауза. Удвоение ДНК перед мейозом происходит в синтетическом периоде интерфазы (как и при митозе).
Помимо типичных для профазы процессов (спирализация ДНК в хромосомы, разрушение ядерной оболочки, движение центриолей к полюсам клетки) в профазе мейоза I происходят два важнейших процесса: конъюгация и кроссинговер.
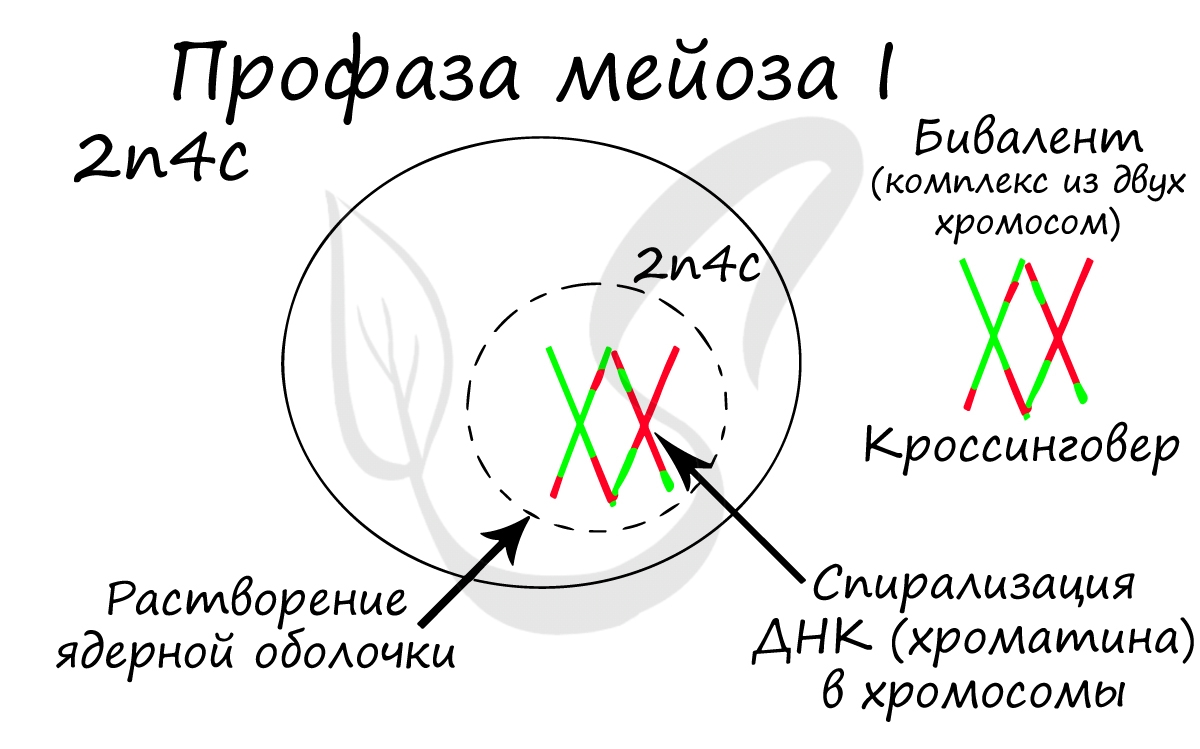
Кроссинговер является важнейшим процессом, в ходе которого возникают рекомбинации генов, что создает уникальный материал для эволюции, последующего естественного отбора. Кроссинговер приводит к генетическому разнообразию потомства.
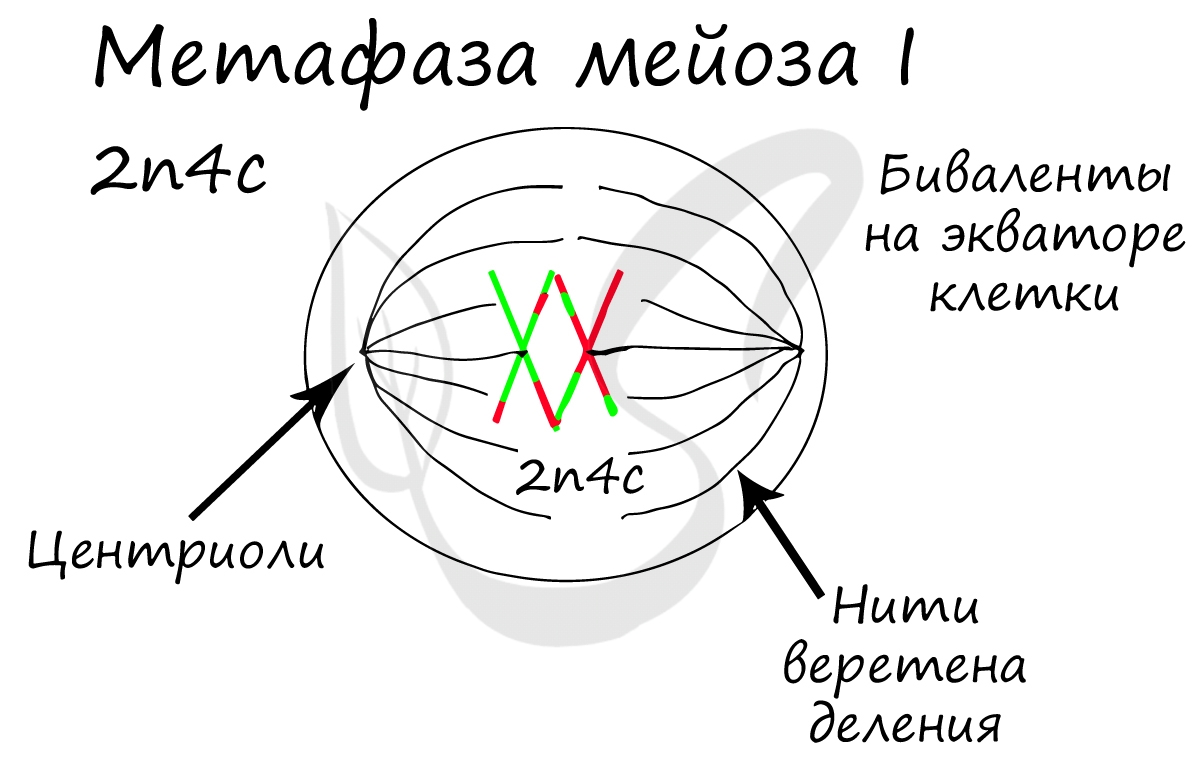
Биваленты (комплексы из двух хромосом) выстраиваются по экватору клетки. Формируется веретено деления, нити которого крепятся к центромере (кинетохору) каждой хромосомы, составляющей бивалент.

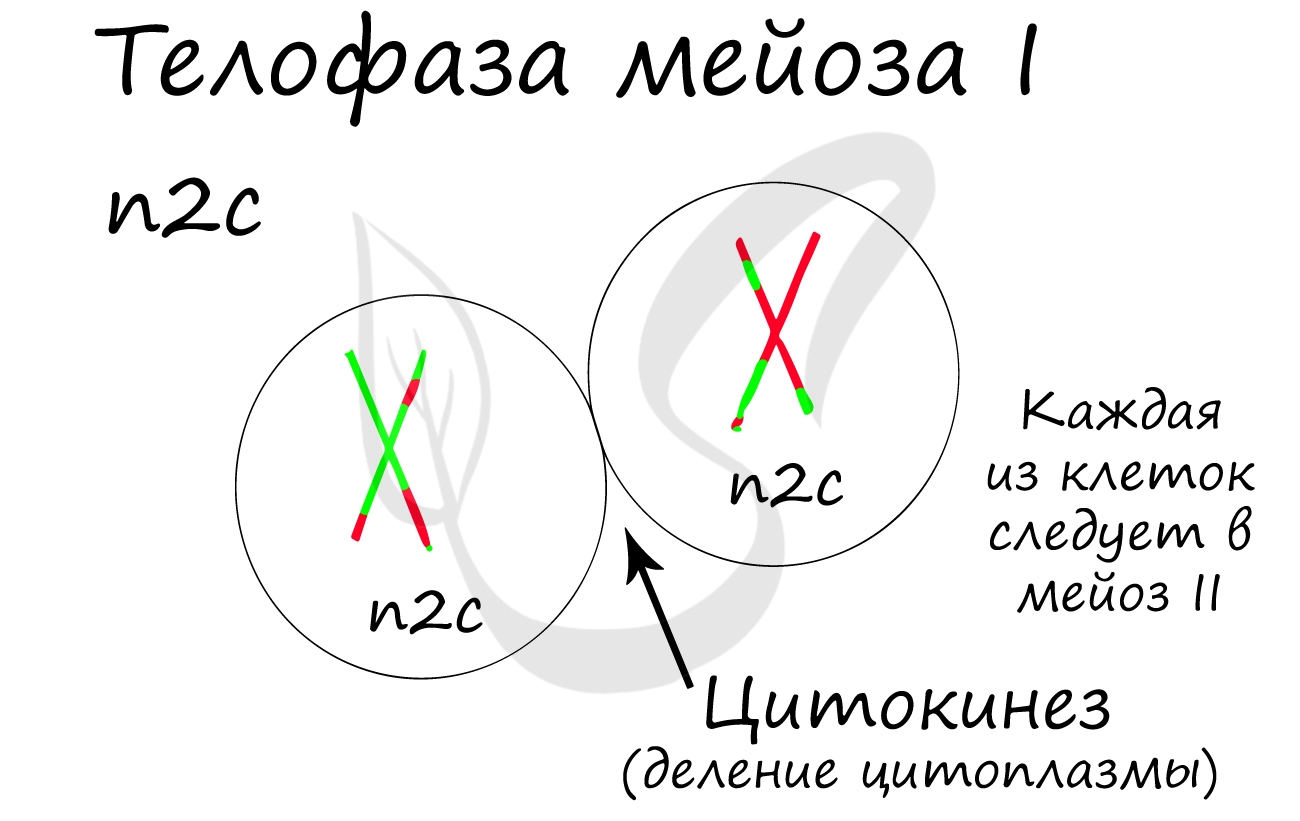
Мейоз II весьма напоминает митоз по всем фазам, поэтому если вы что-то подзабыли: поищите в теме про митоз. Главное отличие мейоза II от мейоза I в том, что в анафазе мейоза II к полюсам клетки расходятся не хромосомы, а хроматиды (дочерние хромосомы).
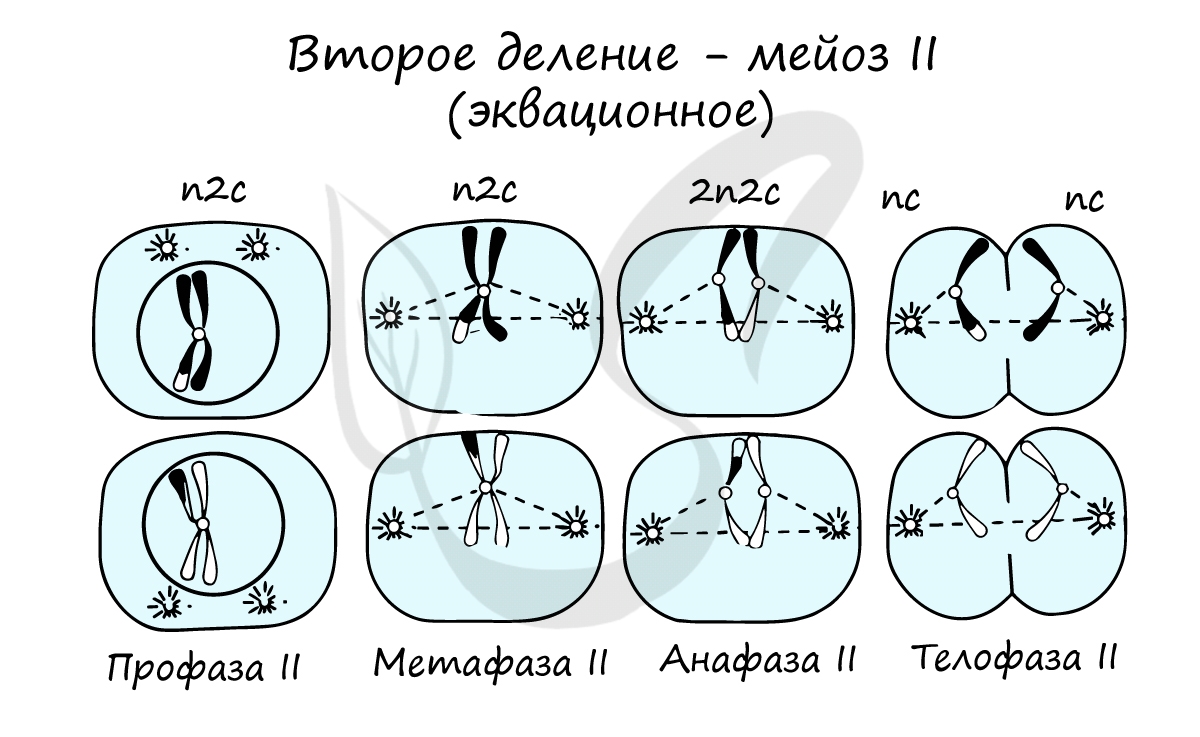
Сейчас мы возьмем клетку, в которой 4 хромосомы. Попытайтесь самостоятельно описать фазы и этапы, через которые она пройдет в ходе мейоза. Проговорите и осмыслите набор хромосом в каждой фазе.
Бинарное деление надвое
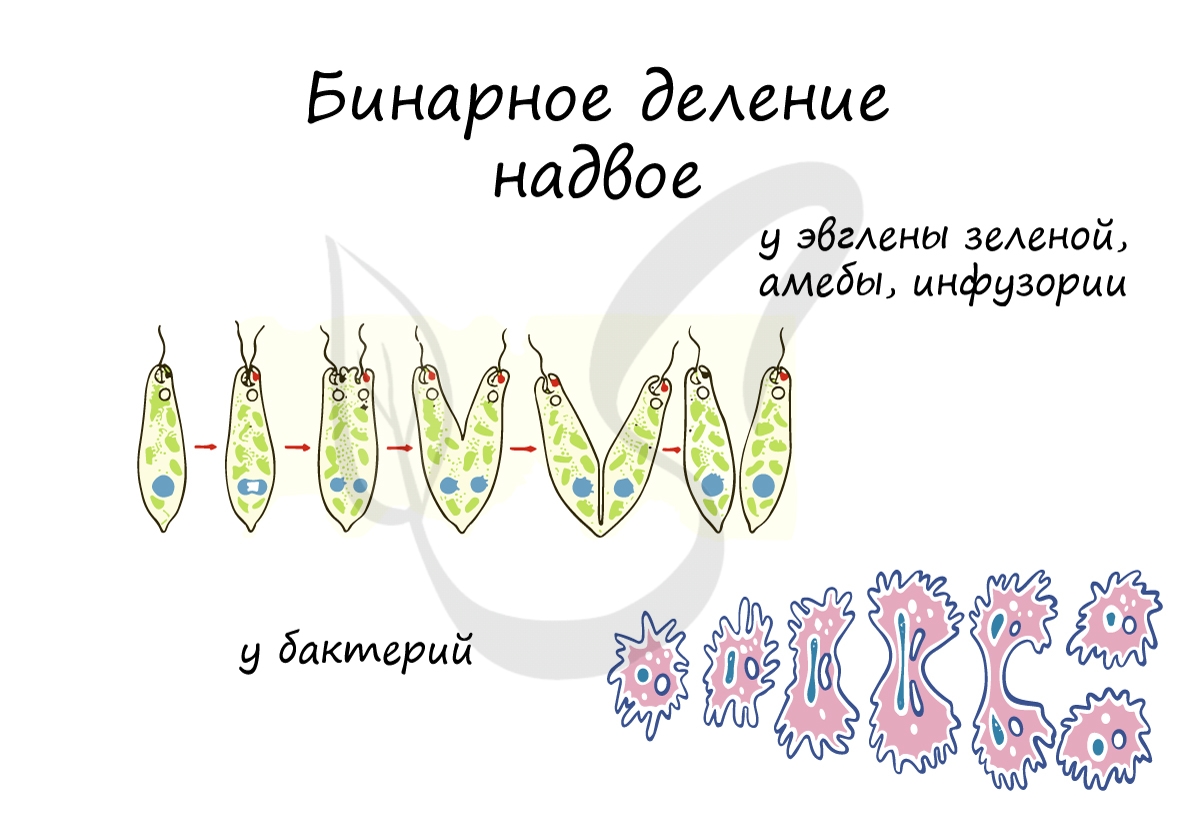
При благоприятных условиях бактерии делятся каждые 20 минут. В случае, если условия не столь благоприятны, то больше времени уходит на рост и развитие, накопление питательных веществ. Интервалы между делениями становятся длиннее.
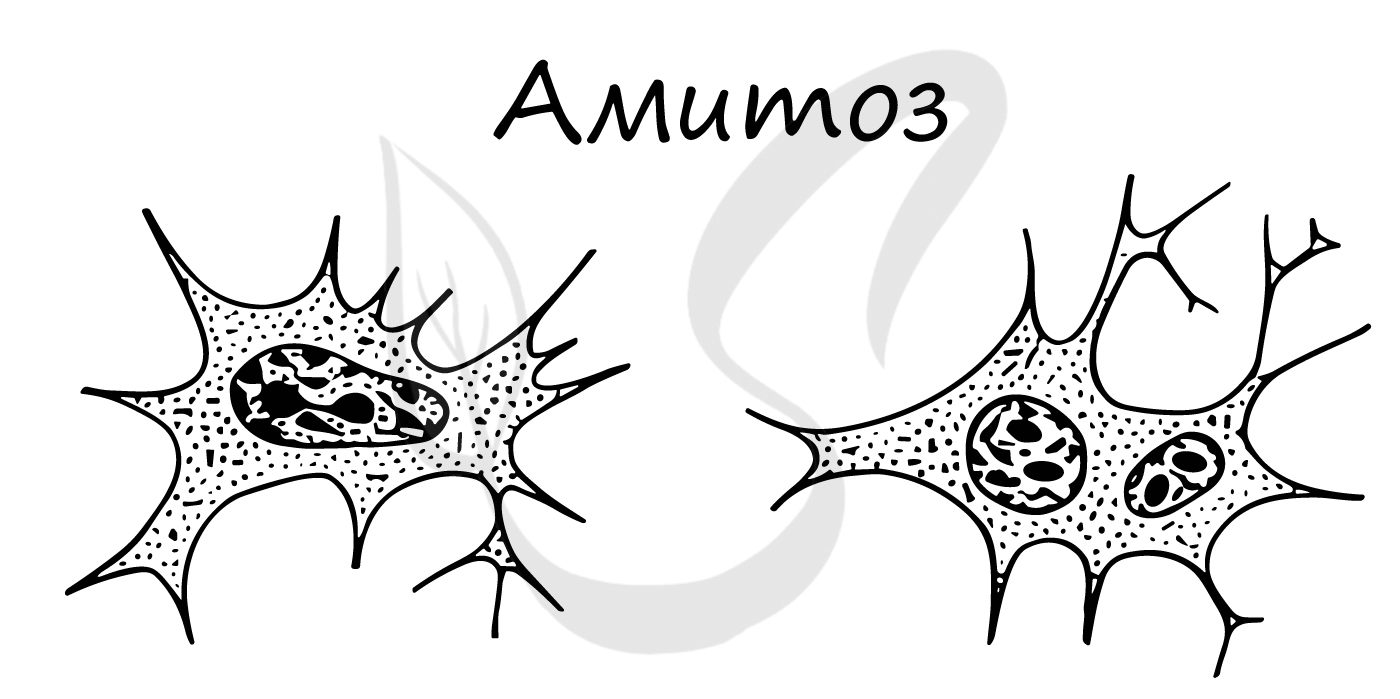
Амитоз встречается в раковых (опухолевых) клетках, воспалительно измененных, в старых клетках.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Тема 4. Сцепление генов. Кроссинговер
Предположения о связи явления наследственности с хромосомами впервые были высказаны еще в конце XIX столетия. Особенно подробно эту идею развивал в своей теории “зародышевой плазмы” А. Вейсман (см. первую лекцию). Позже американский цитолог У. Сэттон обратил внимание на соответствие характера наследования признаков у одного из видов кузнечика поведению хромосом в процессе мейоза. Он сделал вывод о локализации наследственных факторов, определяющих эти признаки, в хромосомах и об ограниченности действия закона независимого комбинирования признаков, установленного Менделем. Он считал, что независимо могут комбинироваться только те признаки, наследственные факторы которых лежат в разных хромосомах. Поскольку количество признаков намного превышает количество пар хромосом, то многие признаки контролируются генами одной хромосомы, которые должны наследоваться совместно.
Первый случай совместного наследования в 1906 г. описали английские генетики У. Бэтсон и Р. Пеннет у душистого горошка (Lathyrus odoratus L.). Они скрестили две расы душистого горошка, различающиеся по двум признакам. Одна раса характеризовалась пурпурной окраской цветков и удлиненной формой пыльцы, другая — красной окраской и округлой формой. Оказалось, что пурпурная окраска полностью доминирует над красной, а удлиненная форма пыльцы над округлой. Каждая пара признаков в отдельности давала расщепление 3 : 1. Гибриды F1 от скрещивания растений этих двух рас наследовали доминантные признаки одного из родителей, т.е. имели пурпурные цветы и удлиненную пыльцу. Однако в F2 соотношение ожидаемых четырех фенотипов не укладывалось в формулу 9 : 3 : 3 : 1, характерную для независимого наследования. Основное отличие заключалось в том, что комбинации признаков, которыми характеризовались родители, встречались чаще, чем следует, в то время как новые сочетания проявлялись в количестве менее ожидаемого. Родительские фенотипы преобладали также в поколении от анализирующего скрещивания. Создавалось впечатление, что наследственные факторы, имевшиеся у родителей, в процессе наследования стремятся оставаться вместе. И, наоборот, факторы, вносимые разными родителями, как бы сопротивляются вхождению в одну гамету. Это явление ученые назвали “притяжением” и “отталкиванием” факторов. При использовании родителей с другими комбинациями этих признаков Бетсон и Пеннет получили такие же результаты.
В течение нескольких лет этот случай необычного наследования у душистого горошка считался отклонением от III закона Менделя. Объяснение ему было дано Т. Морганом и его коллегами, которые обнаружили много случаев подобного наследования признаков у дрозофилы. Согласно их выводам, преимущественная передача потомству исходных комбинаций признаков обусловлена тем, что определяющие их гены располагаются в одной хромосоме, т.е. физически соединены. Это явление было названо Морганом сцеплением генов. Он же дал объяснение неполному сцеплению, предположив, что оно является результатом кроссинговера — перекреста гомологичных хромосом, которые во время коньюгации в профазе мейоза обмениваются гомологичными участками. К такому заключению Морган пришел под влиянием данных голландского цитолога Ф. Янсенса (1909), изучавшего мейоз и обратившего внимание на характерные переплетения хромосом в профазе I, напоминавшие греческую букву c. Он назвал их хиазмами.
Морганом было проделано скрещивание на дрозофиле, которое стало генетическим доказательством наличия обмена генами. В качестве родительских форм он использовал две линии дрозофилы, различающиеся по двум парам признаков. Мухи одной линии имели серое тело (признак дикого типа) и редуцированные крылья (рецессивная мутация vestigal, vg), а мухи другой линии — черное тело (рецессивная мутация black, b) и нормальные крылья. Все гибриды F1 унаследовали доминантные признаки дикого типа — серое тело и нормальные крылья. Далее Морган отступил от обычной схемы скрещивания и вместо F2 получал поколение от скрещивания гибридов F1 с гомозиготными рецессивными особями, т.е. проводил анализирующее скрещивание. Таким способом он пытался точно определить, какие типы гамет и в каком количестве образуют гибриды F1. Были проведены два типа анализирующих скрещиваний: в первом из них гибридные самки скрещивались с гомозиготными рецессивными самцами (bbvgvg), во втором — гомозиготные рецессивные самки скрещивались с гибридными самцами.
Результаты двух анализирующих скрещиваний были разными. Как видно из схемы, Fа прямого скрещивания состоит из четырех фенотипических классов. Это говорит о том, что гибридная самка образует четыре типа гамет, слияние которых с единственной гаметой гомозиготного рецессива приводит к проявлению в Fа четырех разных комбинаций признаков. Два класса, которые повторяют по фенотипу родительские особи, Морган назвал некроссоверными, так как они произошли от слияния гамет, сформированных без участия кроссинговера и обмена генами. По количеству эти классы являются более многочисленными (83%), чем два других класса — кроссоверные (17%), характеризующиеся новыми сочетаниями признаков. Их появление свидетельствовало о том, что в мейозе при образовании части гамет самки идет процесс кроссинговера и осуществляется обмен генами. Такой тип наследования получил название неполного сцепления.
Иные результаты были получены в обратном скрещивании, где анализировался генотип гибридного самца. В Fа в равных количествах были представлены только два класса особей, повторяющие по фенотипу родительские формы. Это говорило о том, что гибридный самец в отличие от гибридной самки формировал с равной частотой гаметы двух типов с исходным сочетанием генов. Подобная ситуация могла иметь место только при условии отсутствия кроссинговера и, следовательно, обмена генами при образовании гамет у самца. Этот тип наследования был назван Морганом полным сцеплением. Позже было установлено, что кроссинговер при образовании гамет у самца, как правило, отсутствует.
Перекрест хромосом происходит в профазе I мейоза, и поэтому его называют мейотическим. Он осуществляется после того как гомологичные хромосомы на стадии зиготены соединяются в пары, образуя биваленты. В профазе I каждая хромосома представлена двумя сестринскими хроматидами, и перекрест происходит не между хромосомами, а между хроматидами гомологов. Кроссинговер можно обнаружить лишь в том случае, если гены находятся в гетерозиготном состоянии (BbVv). При гомозиготном состоянии генов кроссинговер генетически выявить нельзя, так как обмен идентичными генами не дает новых комбинаций на уровне фенотипа.
Коллега Т. Моргана А. Стертевант высказал предположение, что частота кроссинговера зависит от расстояния между генами, и полное сцепление обнаруживают гены, расположенные очень близко друг к другу. На этом основании он предложил использовать этот показатель для определения расстояния между генами. Частота кроссинговера определяется на основе результатов анализирующего скрещивания. Процент кроссинговера рассчитывается как отношение числа кроссоверных особей Fа (т.е. особей с новыми сочетаниями родительских признаков) к общему количеству особей этого потомства (в %). 1% кроссинговера принят за единицу расстояния между генами, которая позже в честь Т. Моргана была названа санти-морганидой (или просто морганидой). Частота перекреста отражает силу сцепления генов: чем меньше частота кроссинговера, тем больше сила сцепления и наоборот.
Исследование явления сцепления генов позволило Моргану сформулировать главную генетическую теорию — хромосомную теорию наследственности. Ее основные положения выглядят следующим образом:
Перекрест, происходящий в одном участке хромосомы, называют одинарным перекрестом. Поскольку хромосома представляет собой линейную структуру значительной протяженности, то в ней одновременно могут происходить несколько перекрестов: двойные, тройные и множественные.
Если кроссинговер идет одновременно в двух соседних участках хромосомы, то частота двойных перекрестов оказывается ниже той, которую можно рассчитать на основании частот одинарных перекрестов. Особенно заметное снижение наблюдается при очень близком расположении генов. В этом случае кроссинговер в одном участке механически препятствует кроссинговеру в другом участке. Это явление получило название интерференции. С увеличением расстояния между генами величина интерференции падает. Эффект интерференции измеряется отношением фактической частоты двойных перекрестов к теоретически ожидаемой их частоте, в случае их полной независимости друг от друга. Это соотношение называется коинциденцией. Фактическая частота двойных перекрестов устанавливается экспериментально в ходе гибридологического анализа по частоте фенотипического класса двойных кроссоверов. Теоретическая частота, согласно закону вероятности, равна произведению частот двух одинарных перекрестов. Например, если в хромосоме имеются три гена а, b и с и кроссинговер между а и b идет с частотой 15%, а между b и с — с частотой 9%, то в случае отсутствия интерференции частота двойного кроссинговера равнялась бы 0,15 x 0,09 = 1,35%. При фактической частоте 0,9%, величина коинциденции выражается отношением и равняется:
| 0,009 | = 0,69 = 69% |
| 0,0135 |
Таким образом, в данном случае из-за интерференции реализовалось только 69% двойных перекрестов.
Среди 8 фенотипических классов, образующихся в Fа при наличии трех пар сцепленных признаков два класса двойных кроссоверов являются самыми малочисленными с учетом явления интерференции и в соответствии с законом вероятности.
Существование множественных перекрестов приводит к увеличению изменчивости гибридного потомства, так как благодаря им возрастает число генных комбинаций и, соответственно, число типов гамет у гибридов.
На определении частот одинарных, двойных, тройных и т.д. перекрестов основан принцип построения генетических карт. Генетическая карта — это схема, отражающая порядок расположения генов в хромосоме. За основу расчета расстояния между генами берется процент одинарного кроссинговера между ними. К нему добавляются поправки на величину двойного и более сложных перекрестов, которые уточняют расчет. Если мы имеем три гена, то порядок их взаиморасположения в хромосоме определяется на основании фенотипа класса двойных кроссоверов. При двойном кроссинговере идет обмен средним геном. Следовательно, признак, по которому двойные кроссоверы отличаются от родительских особей, определяется этим геном. Например, если гомозиготная серая длиннокрылая самка дрозофилы с красными глазами (все признаки дикого типа доминантные) скрещивалась с гомозиготным темным (рецессивная мутация black) самцом с редуцированными крыльями (рецессивная мутация ) и яркими глазами (рецессивная мутация cinnabar), и в Fа самыми малочисленными парными классами (т.е. двойными кроссоверами) были серые мухи с яркими глазами и длинными крыльями и черные с красными глазами и редуцированными крыльями, то, следовательно, ген, контролирующий окраску глаз, является средним. Отрезок карты с этими тремя генами будет выглядеть следующим образом:

На генетической карте любой хромосомы отсчет расстояния начинается с нулевой точки — локуса первого гена — и отмечается не расстояние между двумя соседними генами, а расстояние в морганидах каждого последующего гена от нулевой точки.
Генетические карты составлены только для хорошо изученных в генетическом отношении объектов, как прокариотических, так и эукариотических, таких как, например, фаг l, E. coli, дрозофила, мышь, кукуруза, человек. Они являются плодом огромного и систематического труда многих исследователей. Наличие таких карт позволяет предсказывать характер наследования изучаемых признаков, а при селекционной работе — вести сознательный подбор пар для скрещивания.
Генетические доказательства наличия кроссинговера, полученные в опытах Т. Моргана и его коллег, получили прямое подтверждение на цитологическом уровне в 30-х гг. в работах К. Штерна на дрозофиле и Б. МакКлинток и Г. Крейтона на кукурузе. Им удалось сконструировать гетероморфную пару хромосом (пара Х-хромосом у дрозофилы и IV пара аутосом у кукурузы), в которой гомологи имели различную форму. Обмен участками между ними приводил к образованию разных цитологических типов этой пары хромосом, которые можно было идентифицировать цитологически (под микроскопом). Благодаря генетическому маркированию каждому цитологическому типу бивалента соответствовал определенный фенотипический класс потомства.
В 30-х гг. в слюнных железах дрозофилы Т. Пайнтером были обнаружены гигантские, или политенные, хромосомы. Благодаря своим крупным размерам и четкой структурной организации они стали основным объектом цитогенетических исследований. Каждой хромосоме свойствен специфический рисунок из темных полос (дисков) и светлых промежутков (междисков), соответствующих гетерохроматическим и эухроматическим участкам хромосомы. Постоянство этой внутренней структуры гигантских хромосом дало возможность проверить, насколько порядок генов, установленный на основании определения частоты кроссинговера, отражает действительное расположение генов в хромосоме. С этой целью проводится сравнение структуры нормальной хромосомы и хромосомы, несущей хромосомную мутацию, например выпадение или удвоение участка хромосомы. Такое сравнение полностью подтверждает соответствие порядка расположения генов на генетических картах их расположению на хромосомах. Графическое изображение гигантской хромосомы с указанием локализации генов в определенных ее участках называется цитологической картой.
Явление кроссинговера обнаружено не только в половых клетках, но и в соматических. Обычно гомологичные хромосомы в профазе митоза не коньюгируют и располагаются отдельно друг от друга. Однако еще в 1916 г. исследователям иногда удавалось наблюдать картины синапсиса гомологичных хромосом в митотической профазе с образованием фигур перекреста (хиазм). Это явление получило название соматического, или митотического, кроссинговера. На фенотипическом уровне о нем судят по мозаичному изменению признаков в некоторых участках тела. Так, у самок дрозофилы дикого типа, гетерозиготных по рецессивным мутациям yellow (желтое тело) и singed (опаленные щетинки), в результате соматического перекреста могут появиться пятна с рецессивными признаками. При этом, в зависимости от того, где произойдет перекрест: между указанными выше генами или за их пределами, образуется либо пятно с обоими мутантными признаками, либо с одним из них.
Обычно при кроссинговере идет обмен одинаковыми по размеру гомологичными участками хромосом. Но изредка возможны несимметричные разрывы в хроматидах и обмен неравными участками, т.е. неравный кроссинговер. В результате такого обмена оба аллеля какого-либо гена могут оказаться в одной хромосоме (дупликация), а в другом гомологе возникает его нехватка. Подобное изменение обнаружено в Х-хромосоме дрозофилы в участке, содержащим доминантную мутацию Bar (В), определяющую развитие полосковидных глаз с уменьшенным числом фасеток (у гомозигот 70 вместо 700). Дупликация этого гена в результате неравного кроссинговера приводит к дальнейшей редукции числа фасеток (до 25). Цитологически неравный кроссинговер легко выявляется по изменению рисунка гигантских хромосом.
Перекрест хромосом, как сложный физиологический процесс, подвержен сильному влиянию внешних и внутренних факторов. Большое влияние на частоту кроссинговера оказывает структура хромосомы, в первую очередь наличие в ней крупных блоков гетерохроматина. Установлено, что у дрозофилы кроссинговер редко идет вблизи центромеры и на концах хромосом, что обусловлено присутствием прицентромерного и теломерного гетерохроматина. Плотная спирализация гетерохроматических участков хромосомы уменьшает расстояние между генами и препятствует их обмену. На частоту кроссинговера влияют различные хромосомные перестройки и генные мутации. При наличии в хромосоме нескольких инверсий они могут стать “запирателями” перекреста. У кукурузы обнаружены гены, нарушающие процесс коньюгации и тем самым препятствующие кроссинговеру.
У большинства изученных животных и растений мейотический перекрест осуществляется у обоих полов. Но есть отдельные виды животных, у которых кроссинговер идет только у гомогаметного пола, а у гетерогаметного пола отсутствует. Причем кроссинговер не происходит не только в половых хромосомах, но и в аутосомах. Подобная ситуация наблюдается у самцов дрозофилы и самок шелкопряда с кариотипом ХY. Однако у многих видов млекопитающих, птиц, рыб и насекомых гетерогаметность пола не сказывается на процессе кроссинговера.
На процесс кроссинговера влияет функциональное состояние организма. Установлено, что частота перекреста зависит от возраста, как и уровень аномалий в мейозе. С возрастом происходит снижение активности ферментативных систем, в том числе и тех, которые регулируют процесс обмена участками хромосом.
Частоту перекреста можно повысить или понизить влиянием на организм различных факторов внешней среды, таких как высокая и низкая температура, ионизирующие излучения, дегидратация, изменение концентрации ионов кальция, магния и др. в среде, действием химических агентов и т.п. В частности, установлено, что у дрозофилы частота кроссинговера возрастает с повышением температуры.
В заключение следует отметить, что процесс кроссинговера очень важен с эволюционной точки зрения. Он является механизмом, с помощью которого осуществляется генетическая рекомбинация и создаются новые благоприятные генотипы. Комбинативная изменчивость, наряду с мутационной, является основой для создания новых форм.
Перейти к чтению других тем книги «Генетика и селекция. Теория. Задания. Ответы»:



