Какой органоид является местом окисления глюкозы?
Какой органоид является местом окисления глюкозы.


Органоид который является источником энергии в клетке?
Органоид который является источником энергии в клетке.

Что не является биополимером : целлюлоза или глюкоза?
Что не является биополимером : целлюлоза или глюкоза?

Какую роль играет процесс окисления глюкозы в организме человека?
Какую роль играет процесс окисления глюкозы в организме человека.

Процессы анаэробного окисления глюкозы протекают в :
Какой органоид является источником энергии?
Какой органоид является источником энергии?

Каков энергетический выход при полном окислении 18 молекул глюкозы?
Каков энергетический выход при полном окислении 18 молекул глюкозы.
Почему цитоплазма не является органоидом?
Почему цитоплазма не является органоидом?

Установите соответствие между органоидами и протекающими в них химическими процессами органоиды А?
Установите соответствие между органоидами и протекающими в них химическими процессами органоиды А.
Хлоропласты процессы 1)образование атф 2)образование со2 3)образование над.
Н 4)образование надф.
Н 5)фотолиз воды 6)превращение атф в адф 7)окисление над.
Н 9)окисление органических кислот 10)окислительное фосфорилирование 11)синтез глюкозы.
Где происходит аэробное окисление глюкозы?
Где происходит аэробное окисление глюкозы?

1 билет 1)? 2)спиной мозг имеет форму бабочки 3)тугая повязка, если имеется то кровьостонавлившие препораты 2 билет 1) в состав клетки входит 80 элементов периодической системы 2).
Пшеница имеет простые листья, жилкование параллельное.
Органоиды клетки
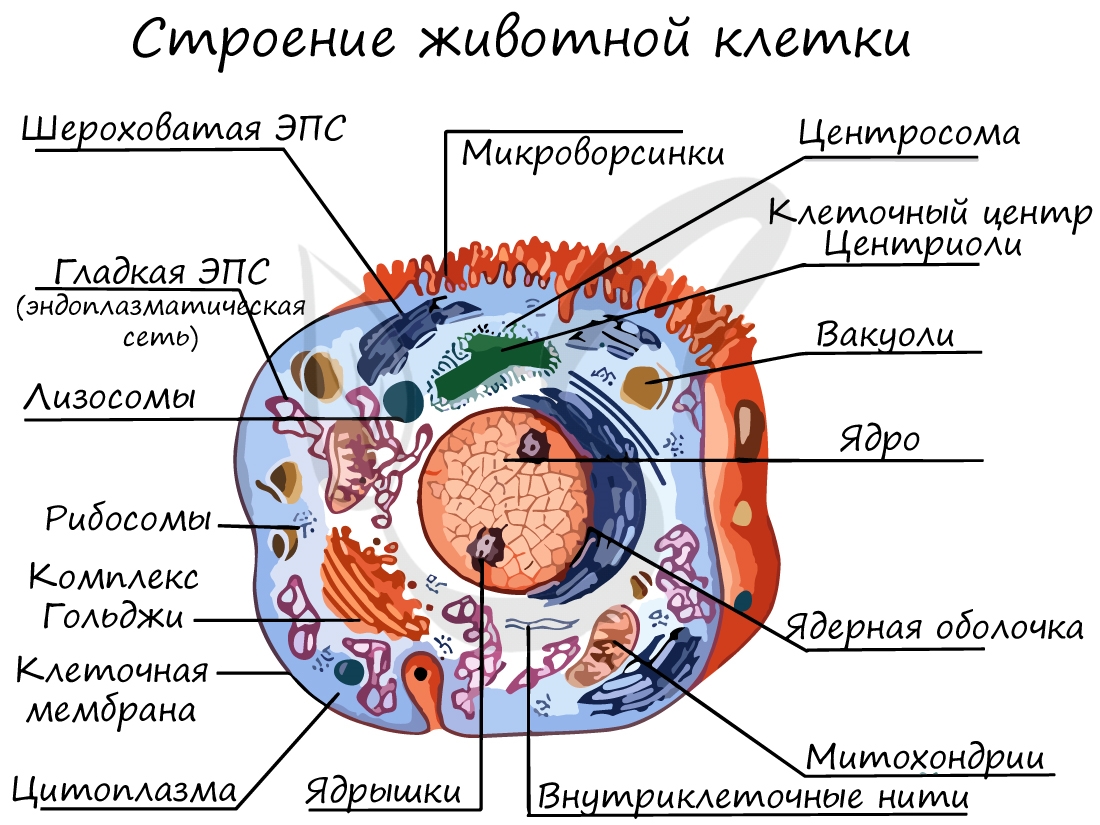
Клеточная мембрана (оболочка)
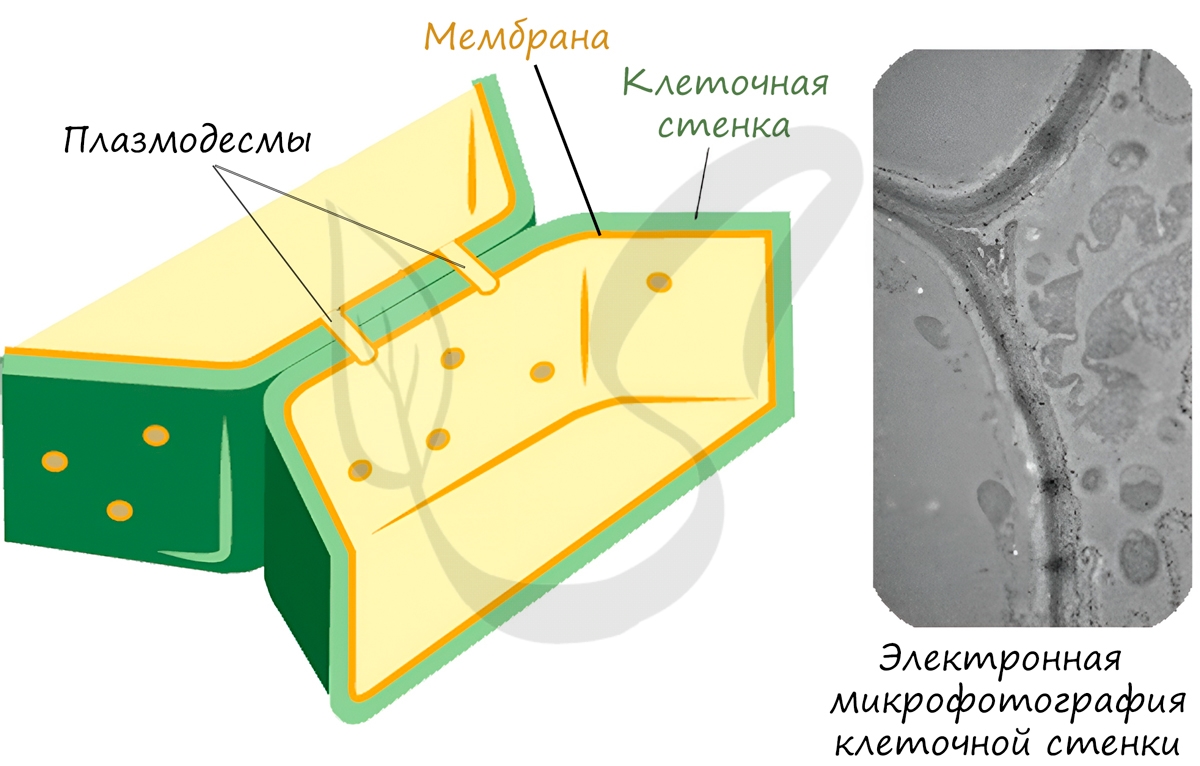
Запомните, что в отличие от клеточной стенки, которая есть только у растительных клеток и у клеток грибов (она придает им плотную, жесткую форму) клеточная мембрана есть у всех клеток без исключения! Этот важный момент объясню еще раз 🙂 У клеток животных имеется только клеточная мембрана, а у клеток растений и грибов есть и клеточная стенка, и клеточная мембрана.
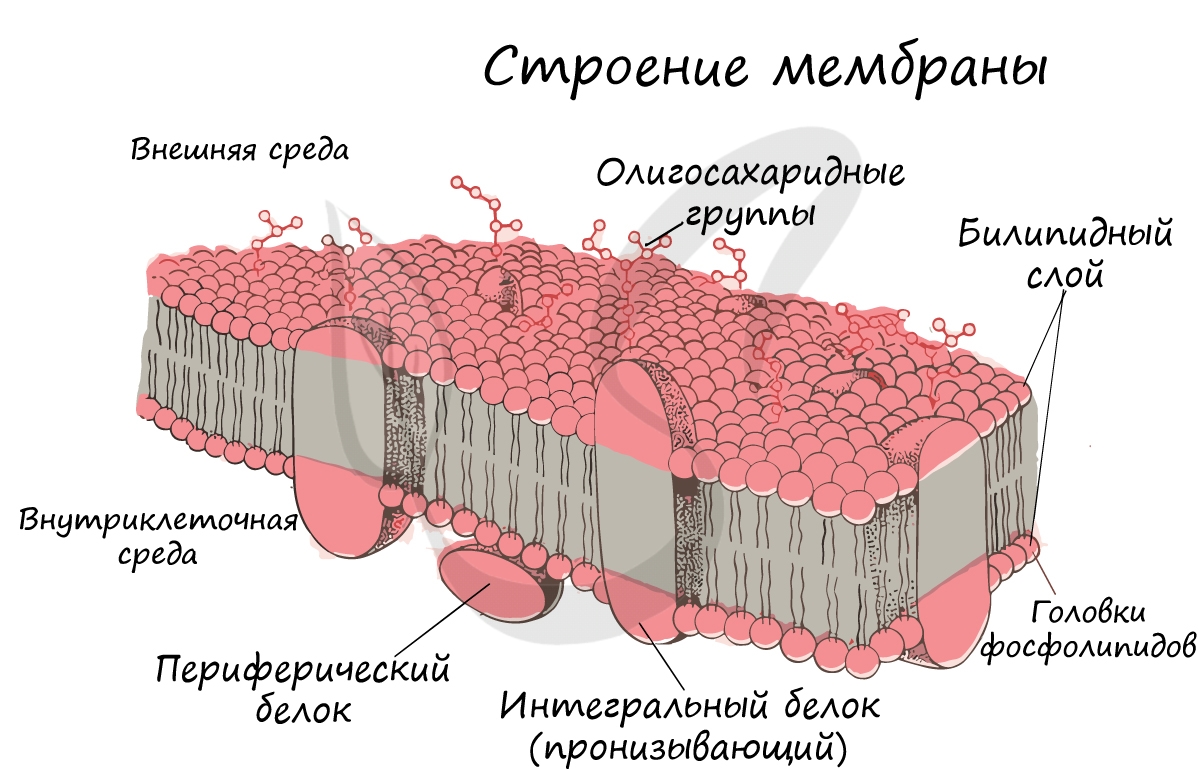
Интегральные (пронизывающие) белки образуют каналы, по которым молекулы различных веществ могут поступать в клетку или удаляться из нее. «Заякоренные» молекулы олигосахаридов на поверхности клетки образуют гликокаликс, который выполняет рецепторную функцию, участвует в избирательном транспорте веществ через мембрану.
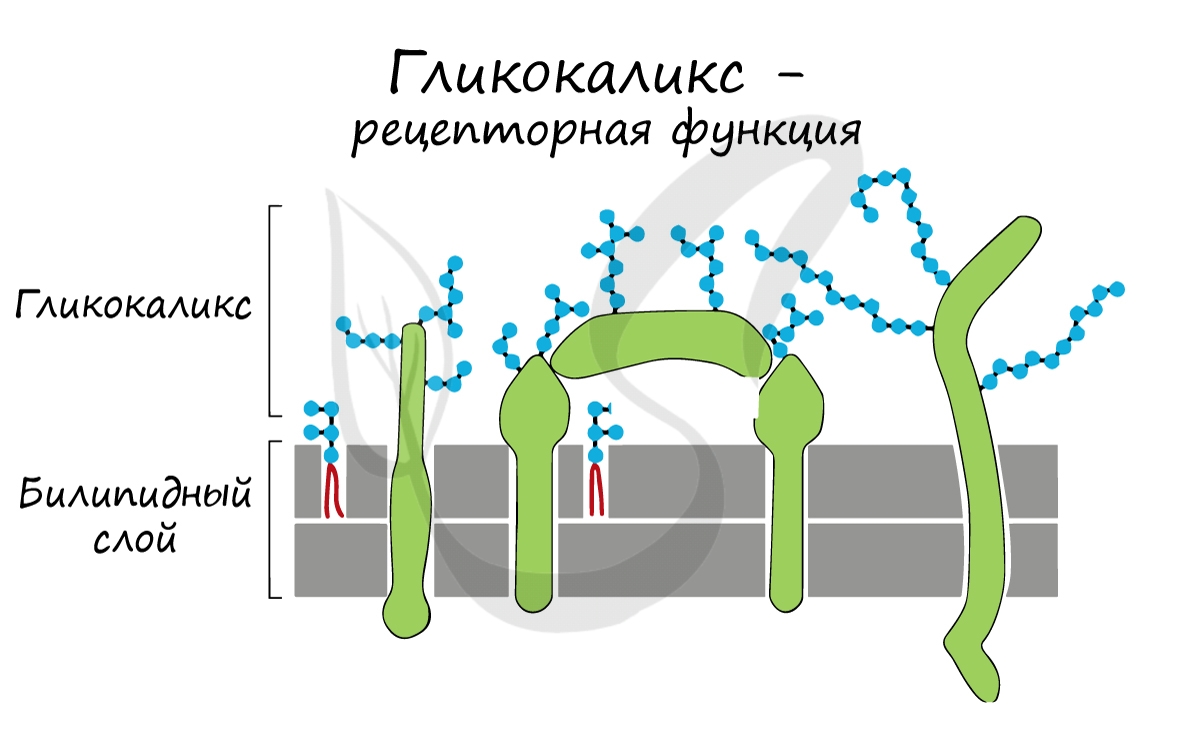
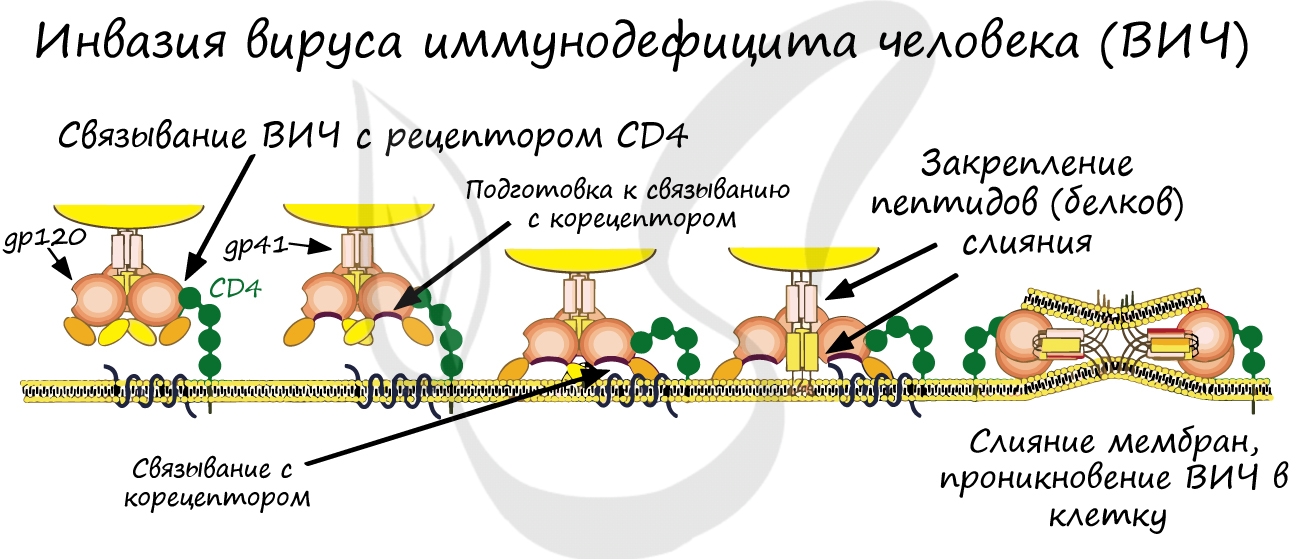
Вирусы и бактерии не являются исключением: они взаимодействуют только с теми клетками, на которых есть подходящие к ним рецепторы. Так, вирус гриппа поражает преимущественно клетки слизистой верхних дыхательных путей. Однако, если рецепторов нет, то вирус не может проникнуть в клетку, и организм приобретает невосприимчивость к инфекции. Вспомните врожденный иммунитет: именно по причине отсутствия рецепторов человек не восприимчив ко многим болезням животных.
Итак, вернемся к клеточной мембране. Ее можно сравнить со стенами помещения, в котором, вероятно, вы находитесь. Стены дома защищают его от ветра, дождя, снега и прочих факторов внешней среды. Рискну предположить, что в вашем доме есть окна и двери, которые по мере необходимости открываются и закрываются 🙂 Так и клеточная мембрана может сообщать внутреннюю среду клетки с внешней средой: через мембрану вещества поступают в клетку и удаляются из нее.
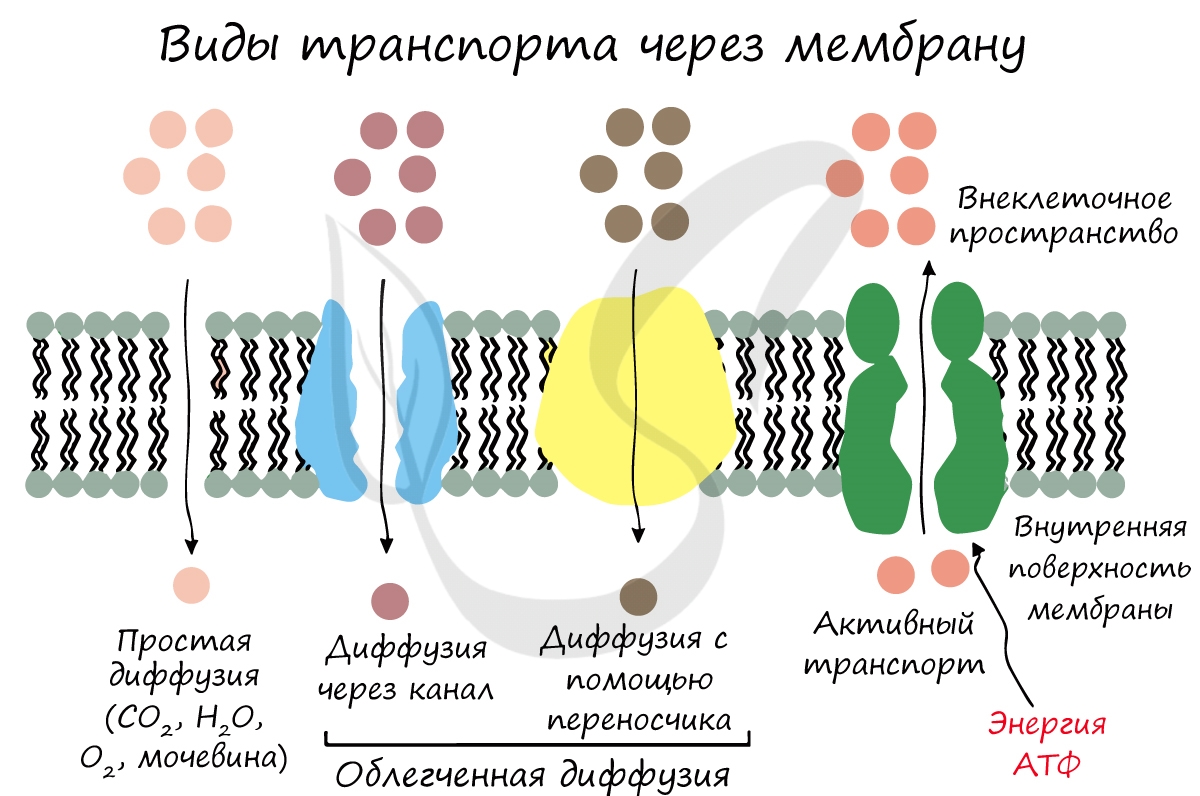
Внутрь клетки с помощью осмоса поступает вода. Путем простой диффузии в клетку попадают O2, H2O, CO2, мочевина. Облегченная диффузия характерна для транспорта глюкозы, аминокислот.
Активный транспорт чаще происходит против градиента концентрации, в ходе него используются белки-переносчики и энергия АТФ. Ярким примером является натрий-калиевый насос, который накачивает ионы калия внутрь клетки, а ионы натрия выводит наружу. Это происходит против градиента концентрации, поэтому без затрат энергии (АТФ) не обойтись.
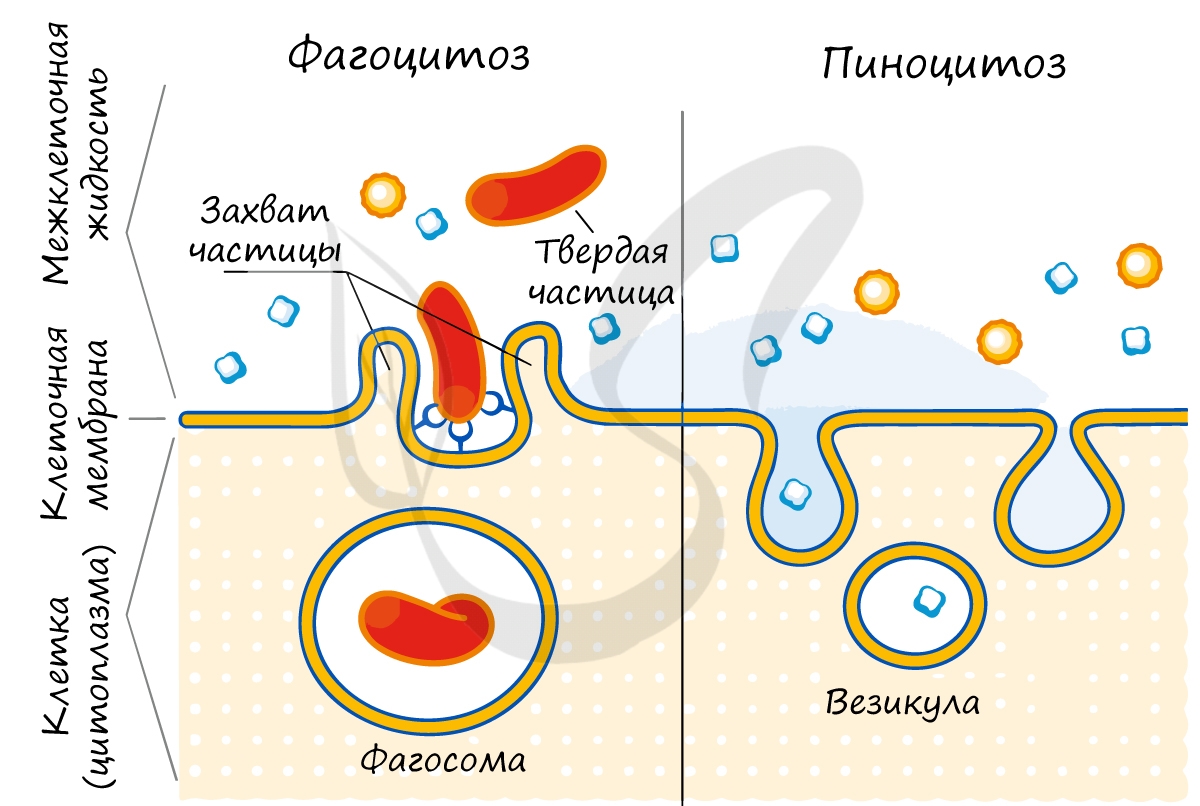
Фагоцитоз был открыт И.И. Мечниковым, который создал фагоцитарную теорию иммунитета. Это теория гласит, что в основе иммунной системы нашего организма лежит явление фагоцитоза: попавшие в организм бактерии уничтожаются фагоцитами (T-лимфоцитами), которые переваривают их.
В ходе эндоцитоза мембрана сильно прогибается внутрь клетки, ее края смыкаются, захватывая бактерию, пищевые частицы или жидкость внутрь клетки. Образуется везикула (пузырек), который движется к пищеварительной вакуоли или лизосоме, где происходит внутриклеточное пищеварение.
Клеточная стенка
Цитоплазма
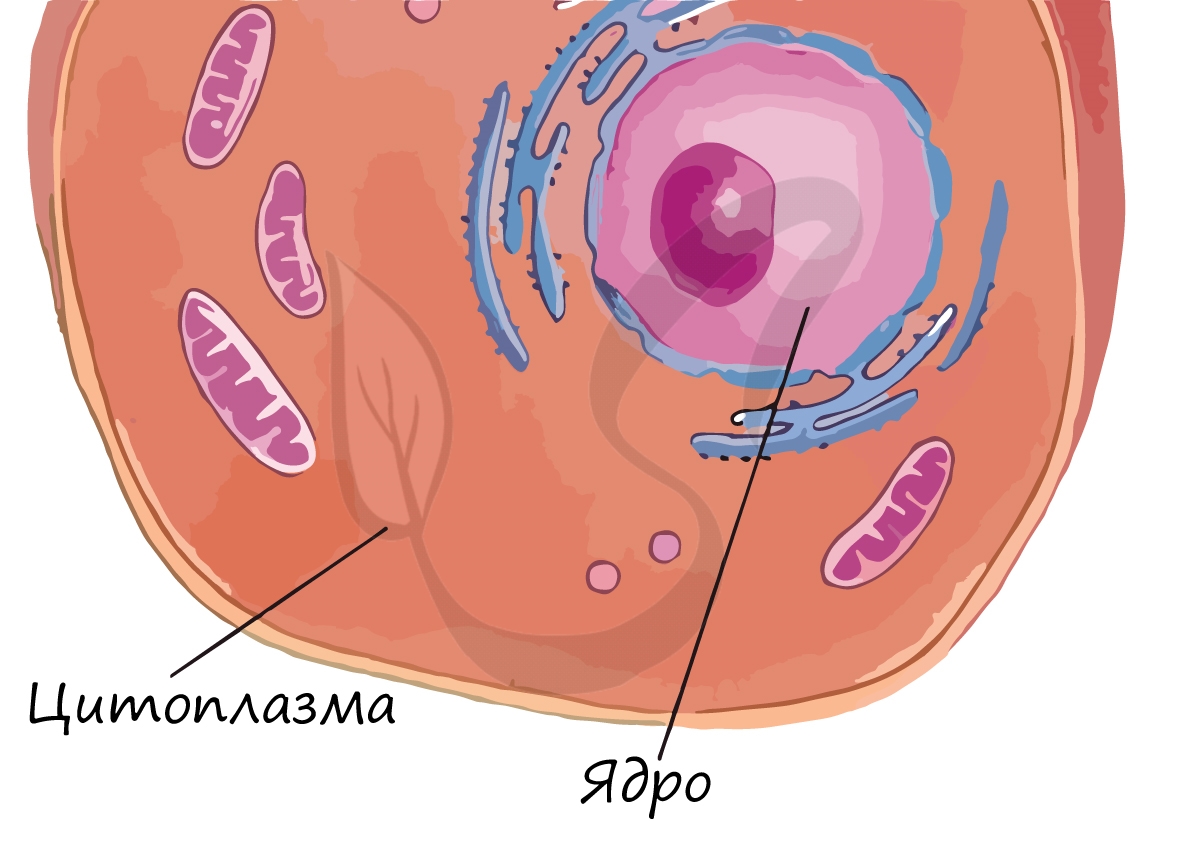
Постоянное движение цитоплазмы поддерживает связь между органоидами клетки и обеспечивает ее целостность.
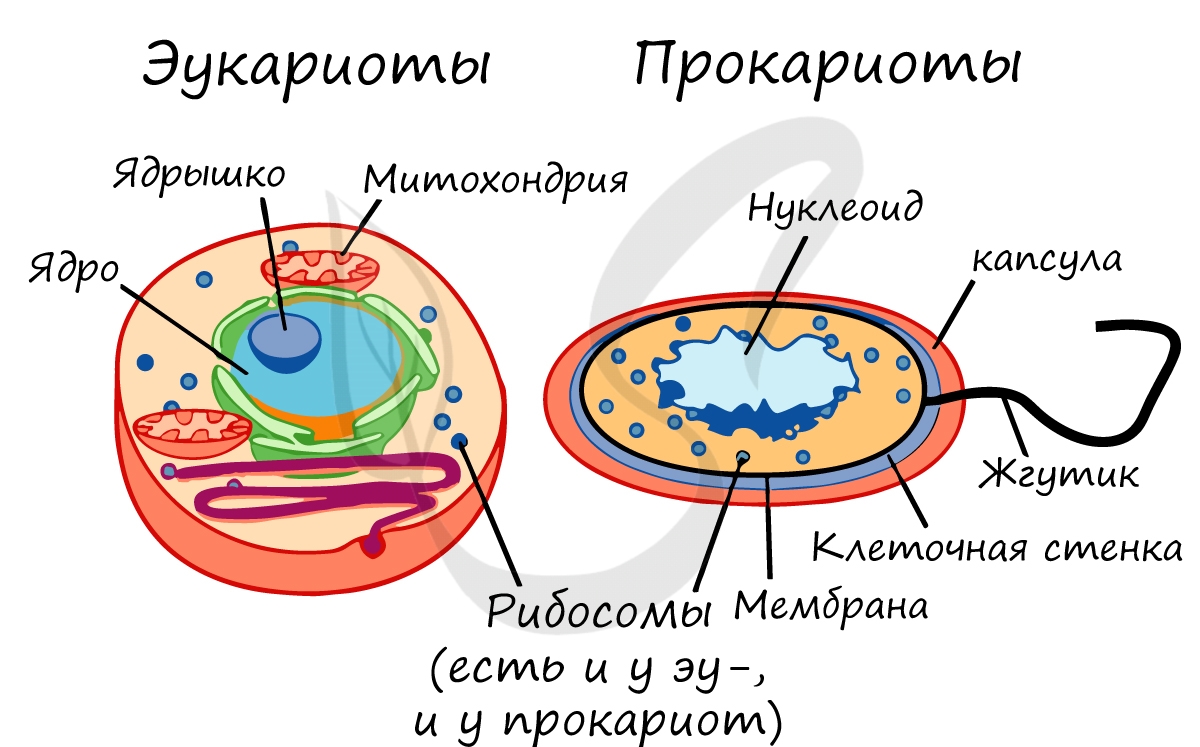
Прокариоты и эукариоты
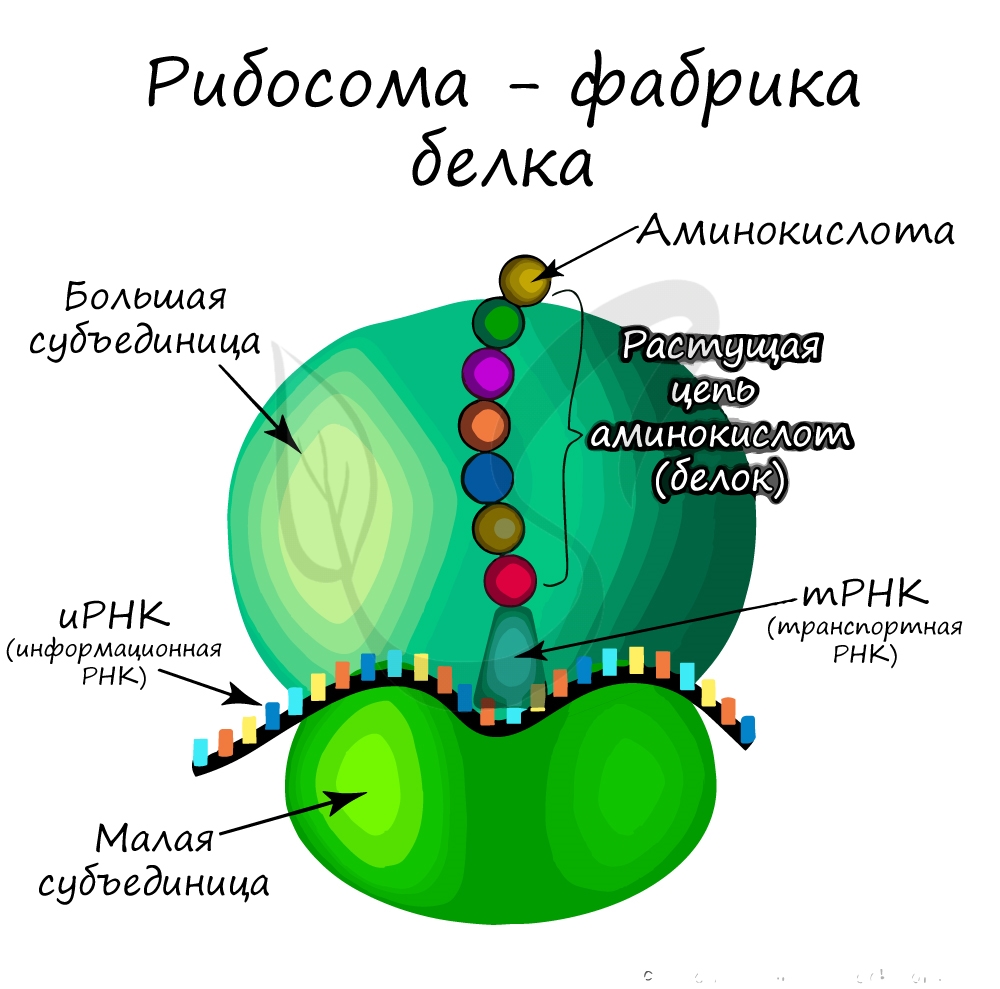
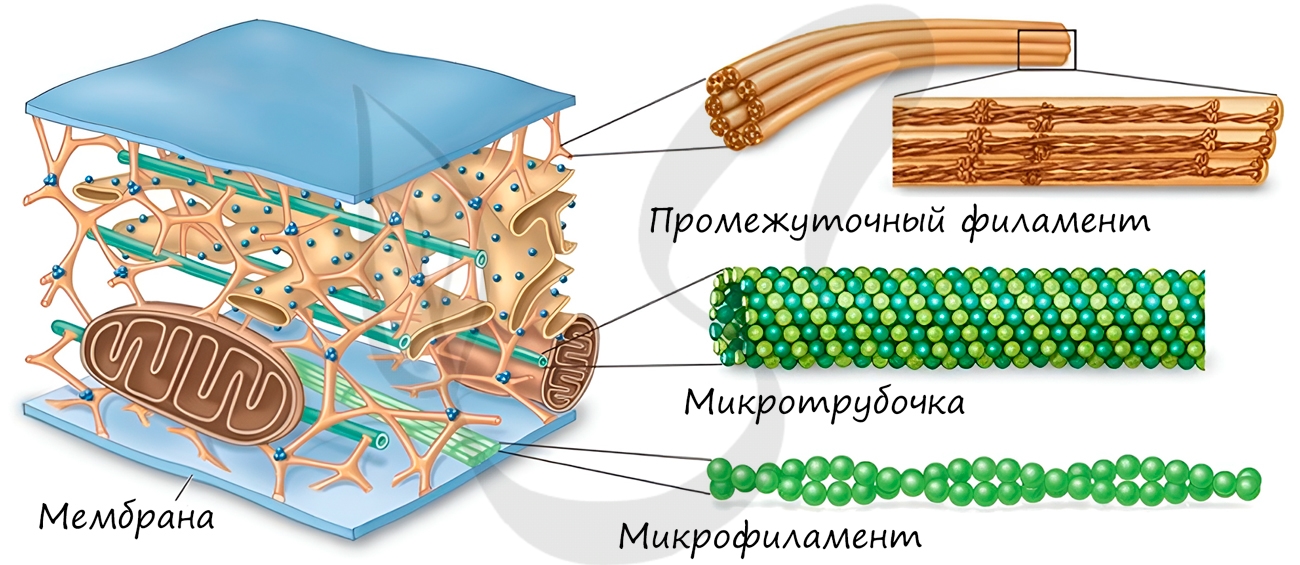
Немембранные органоиды
Очень мелкая органелла (около 20 нм), которая была открыта после появления электронного микроскопа. Состоит из двух субъединиц: большой и малой, в состав которых входят белки и рРНК (рибосомальная РНК), синтезируемая в ядрышке.
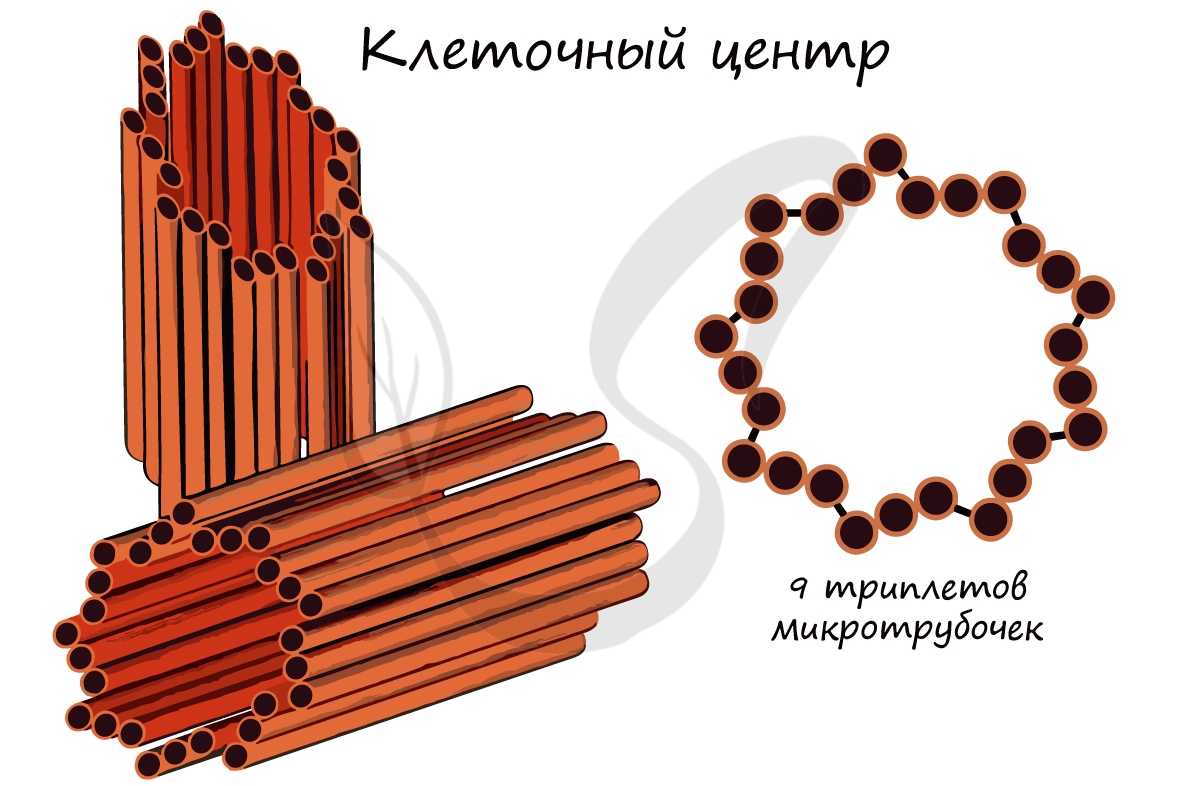
Это органоиды движения, которые выступают над поверхностью клетки и имеют в основе пучок микротрубочек. Реснички встречаются только в клетках животных, жгутики можно обнаружить у животных, растений и бактерий.
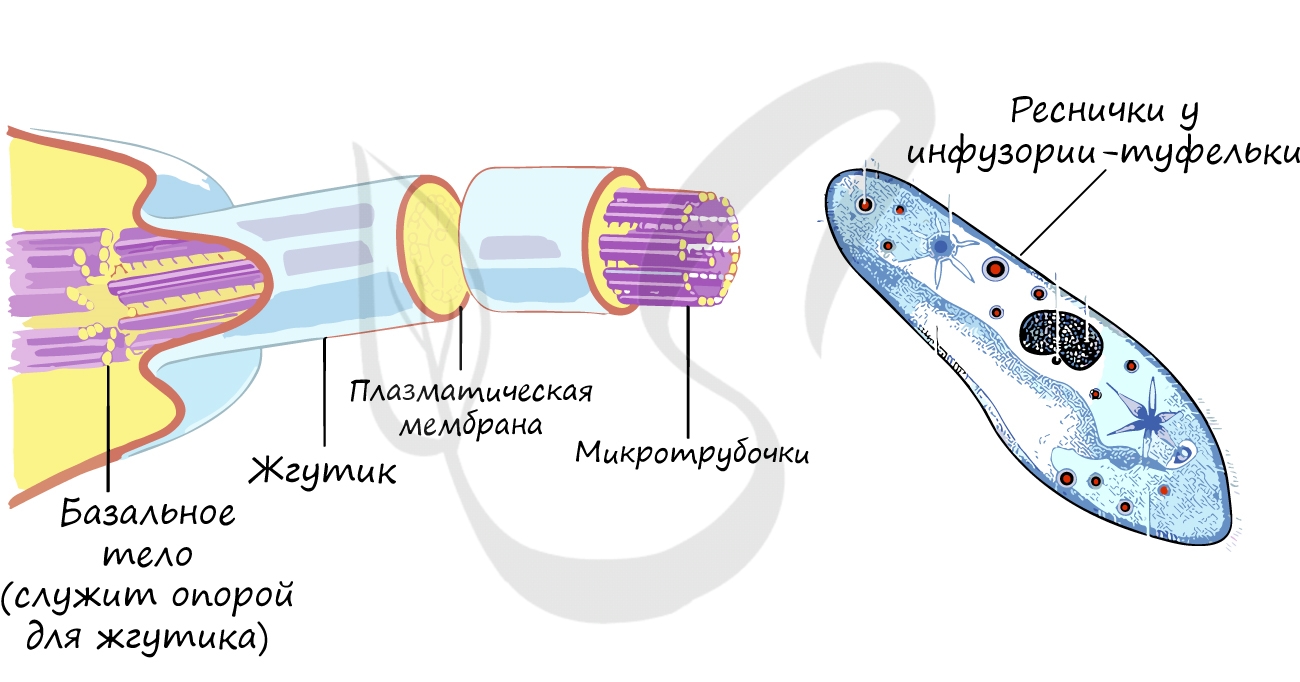
Одномембранные органоиды
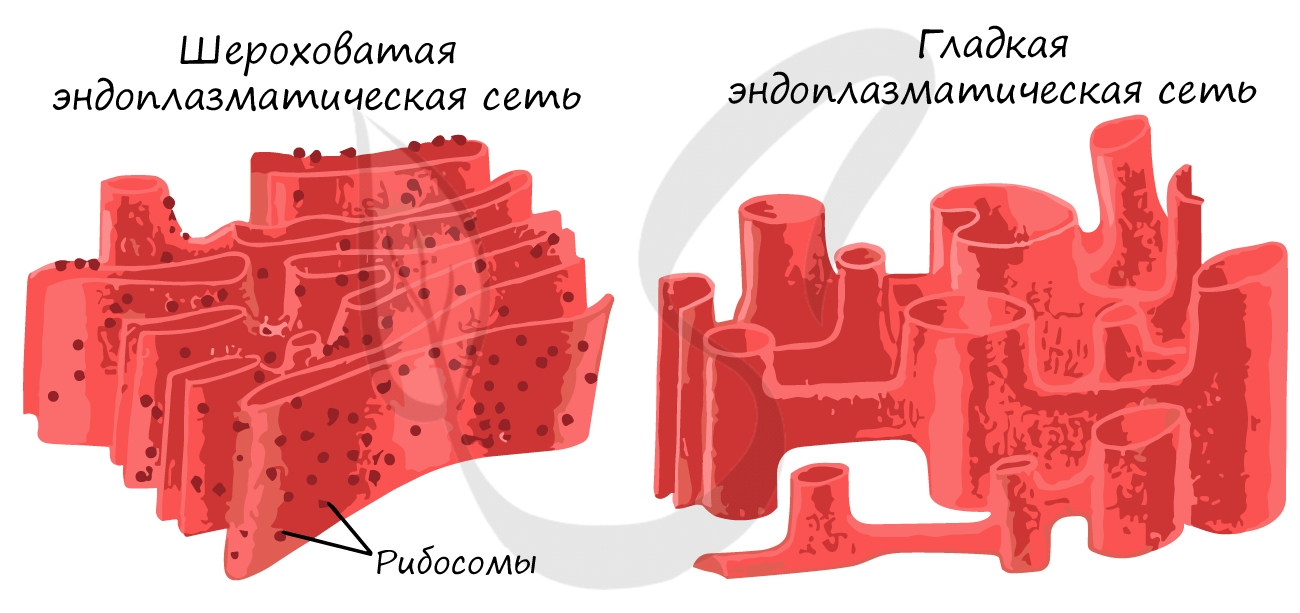
ЭПС представляет собой систему мембран, пронизывающих всю клетку и разделяющих ее на отдельные изолированные части (компартменты). Это крайне важно, так как в разных частях клетки идут реакции, которые могут помешать друг другу, что нарушит процессы жизнедеятельности.
Выделяют гладкую ЭПС и шероховатую ЭПС. Обе они выполняют функцию внутриклеточного транспорта веществ, однако между ними имеются различия. На мембранах гладкой ЭПС происходит синтез липидов, обезвреживаются вредные вещества. Шероховатая ЭПС синтезирует белок, так как имеет на мембранах многочисленные рибосомы (потому и называется шероховатой).
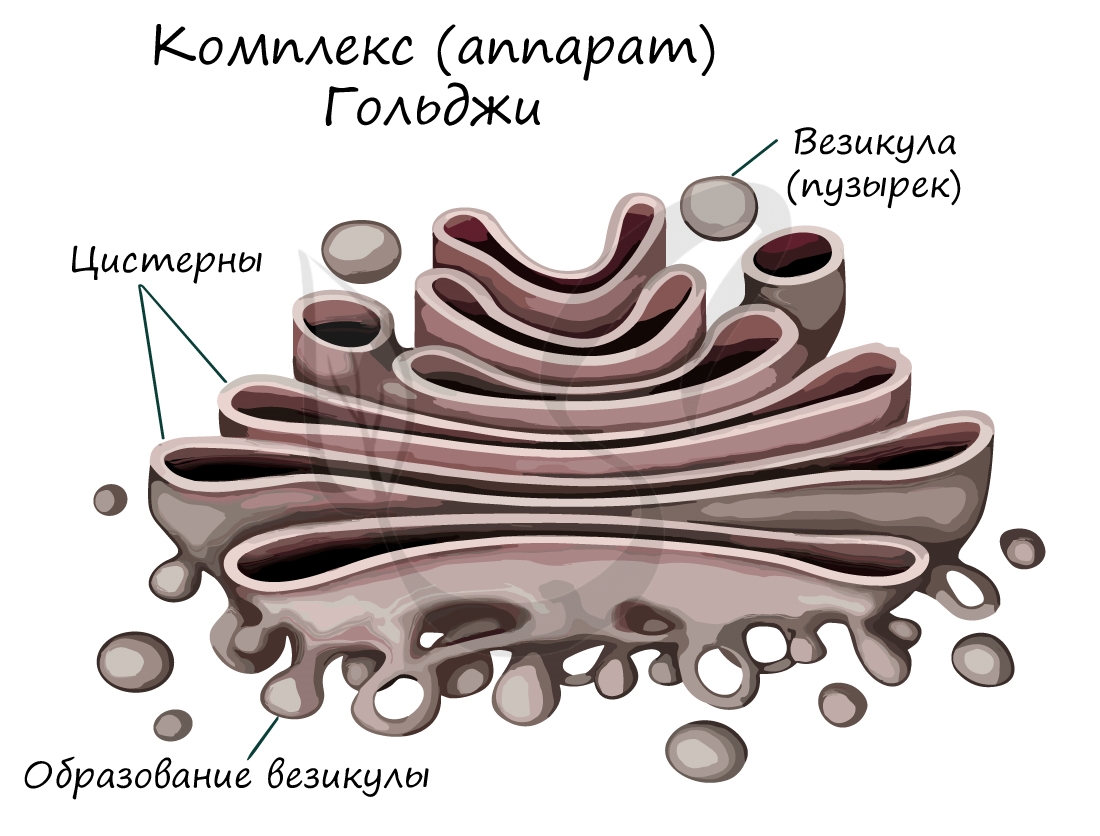
Модифицированные вещества упаковываются в пузырьки и могут перемещаться к мембране клетки, соединяясь с ней, они изливают свое содержимое во внешнюю среду. Можно догадаться, что комплекс Гольджи хорошо развит в клетках эндокринных желез, которые в большом количестве синтезируют и выделяют в кровь гормоны.
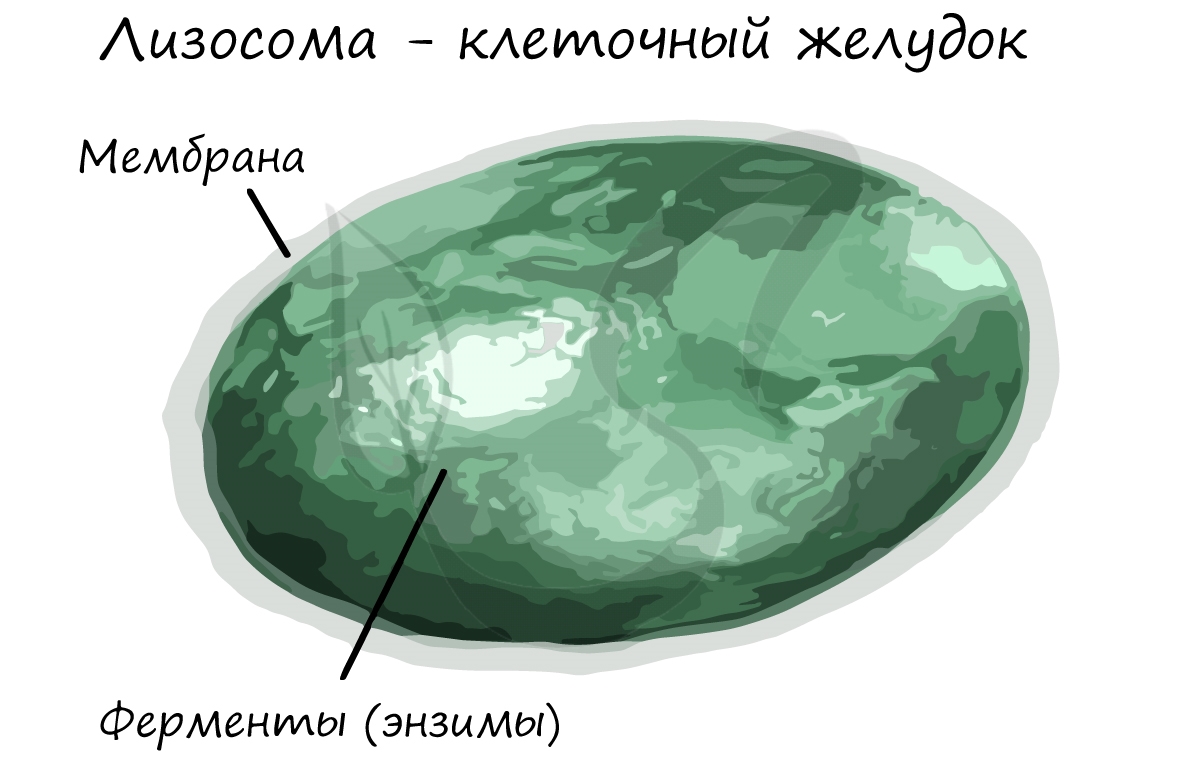
В комплексе Гольджи появляются первичные лизосомы, которые содержат ферменты в неактивном состоянии.

В ходе апоптоза ферменты лизосомы изливаются внутрь клетки, ее содержимое переваривается. Предполагают, что нарушение апоптоза в раковых клетках ведет к бесконтрольному росту опухоли.
Пероксисомы (микротельца) содержат окислительно-восстановительные ферменты, которые разлагают H2O2 (пероксид водорода) на воду и кислород. Если бы пероксид водорода оставался неразрушенными, это приводило бы к серьезным повреждениям клетки.
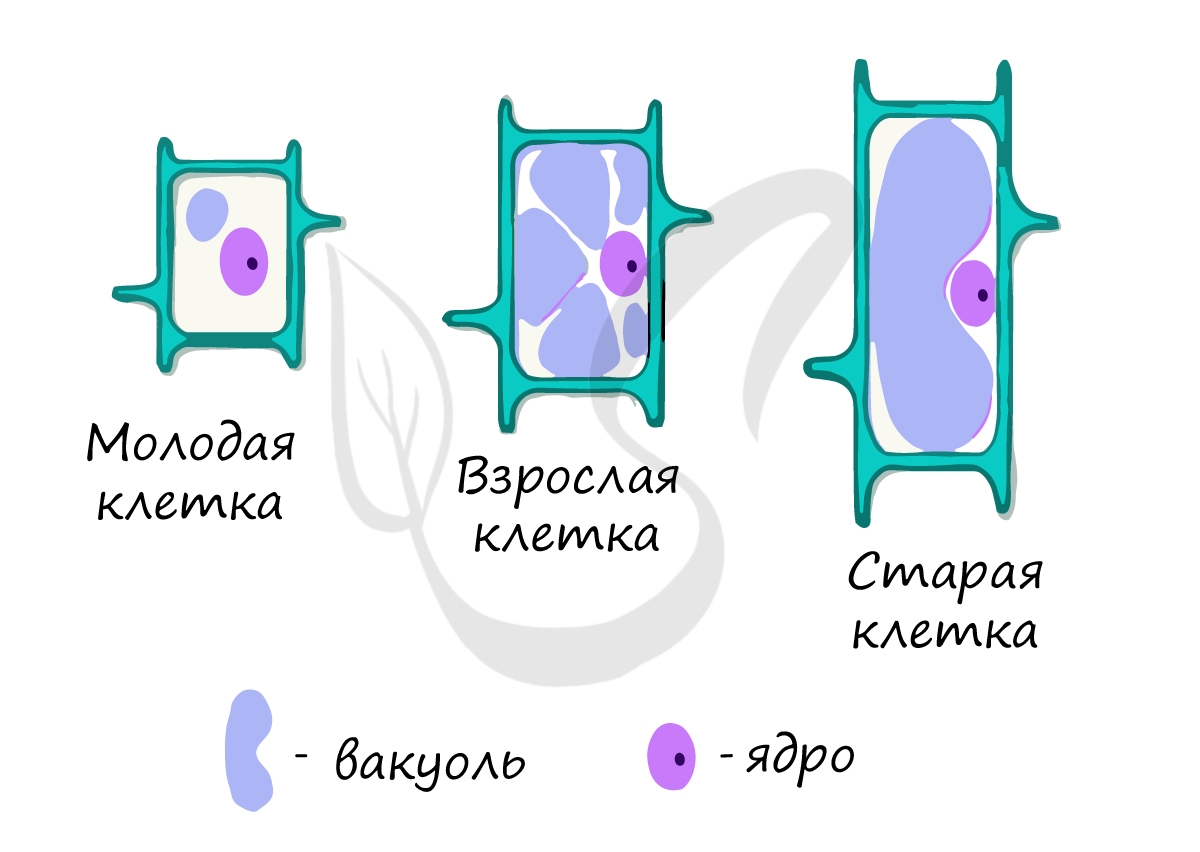
Трудно переоценить значение вакуолей в жизнедеятельности растительной клетки. Вакуоли создают осмотическое давление, придают клетке форму.
Примечательно, что по размеру вакуолей можно судить о возрасте клетки: молодые клетки имеют вакуоли небольшого размера, а в старых клетках вакуоли могут настолько увеличиваться, что оттесняют ядро и остальные органоиды на периферию.
Двумембранные органоиды
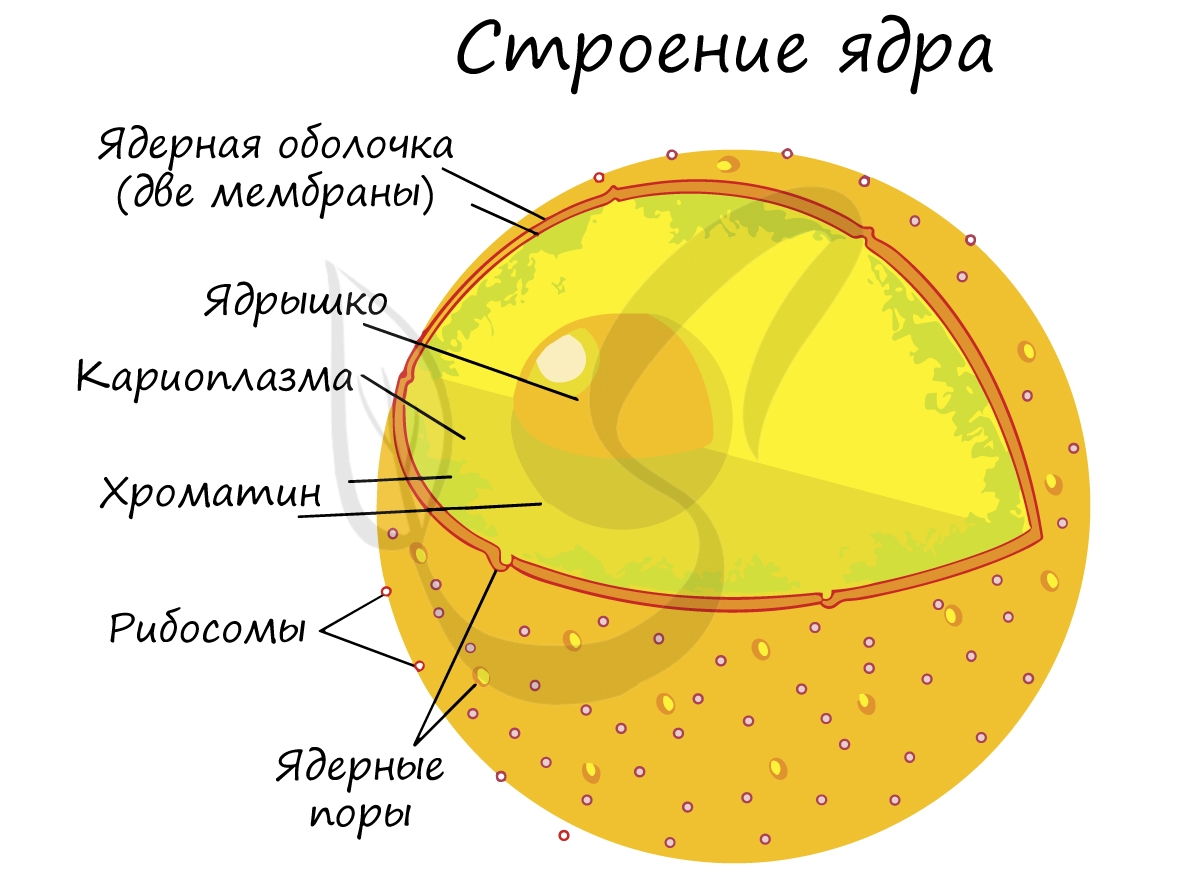
Оболочка ядра состоит из двух мембран и пронизана большим количеством ядерных пор, через которые происходит сообщение между кариоплазмой и цитоплазмой. Главными функциями ядра является хранение, защита и передача наследственного материала дочерним клеткам.
Замечу, что хромосомы видны только в момент деления клетки. Хромосомы представляют собой сильно спирализованные молекулы ДНК, связанные с белками.
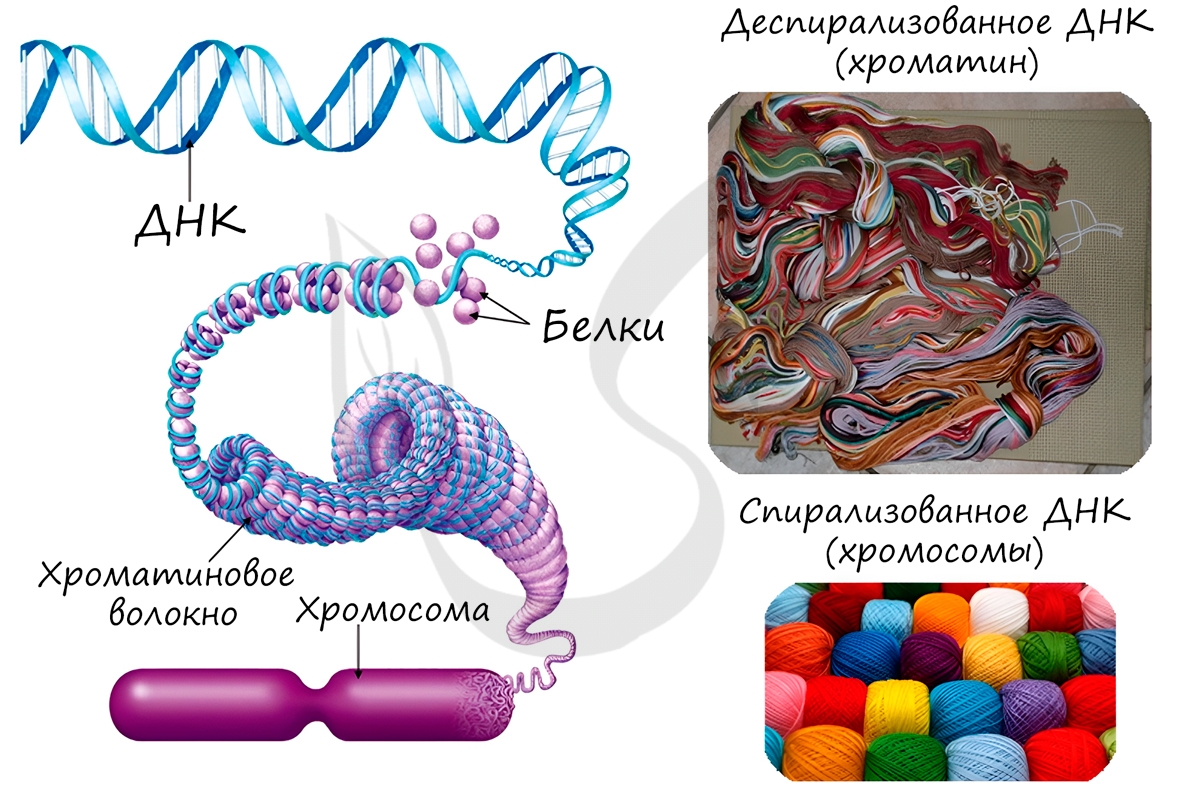
Хромосомы отличаются друг от друга по строению, форме, размерам. Совокупность всех признаков (форма, число, размер) хромосом называется кариотип. Кариотип может быть представлен по-разному: существует кариотип вида, особи, клетки.
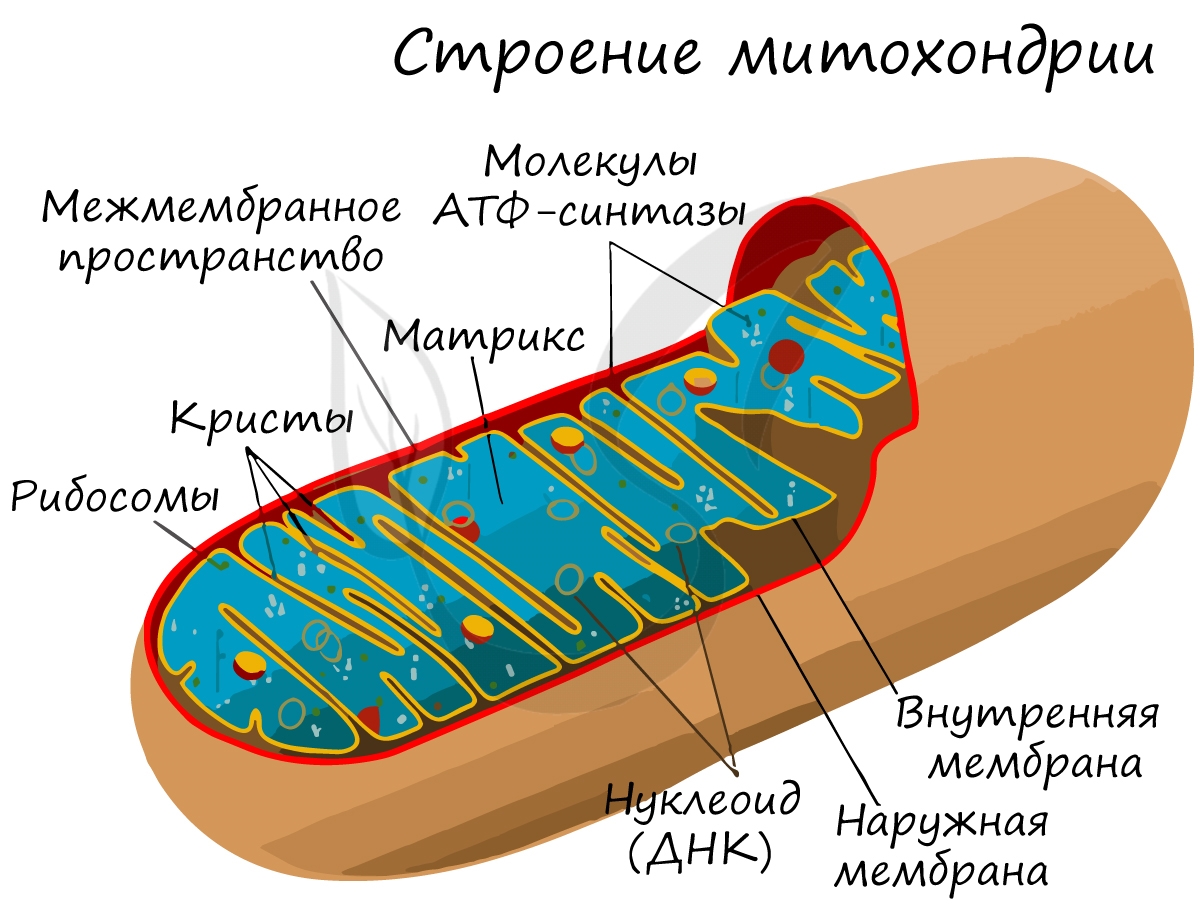
В связи с этим, митохондрия считается полуавтономным органоидом. Вероятнее всего, изначально митохондрии были самостоятельными организмами, однако со временем вступили в симбиоз с эукариотами и стали частью клетки.
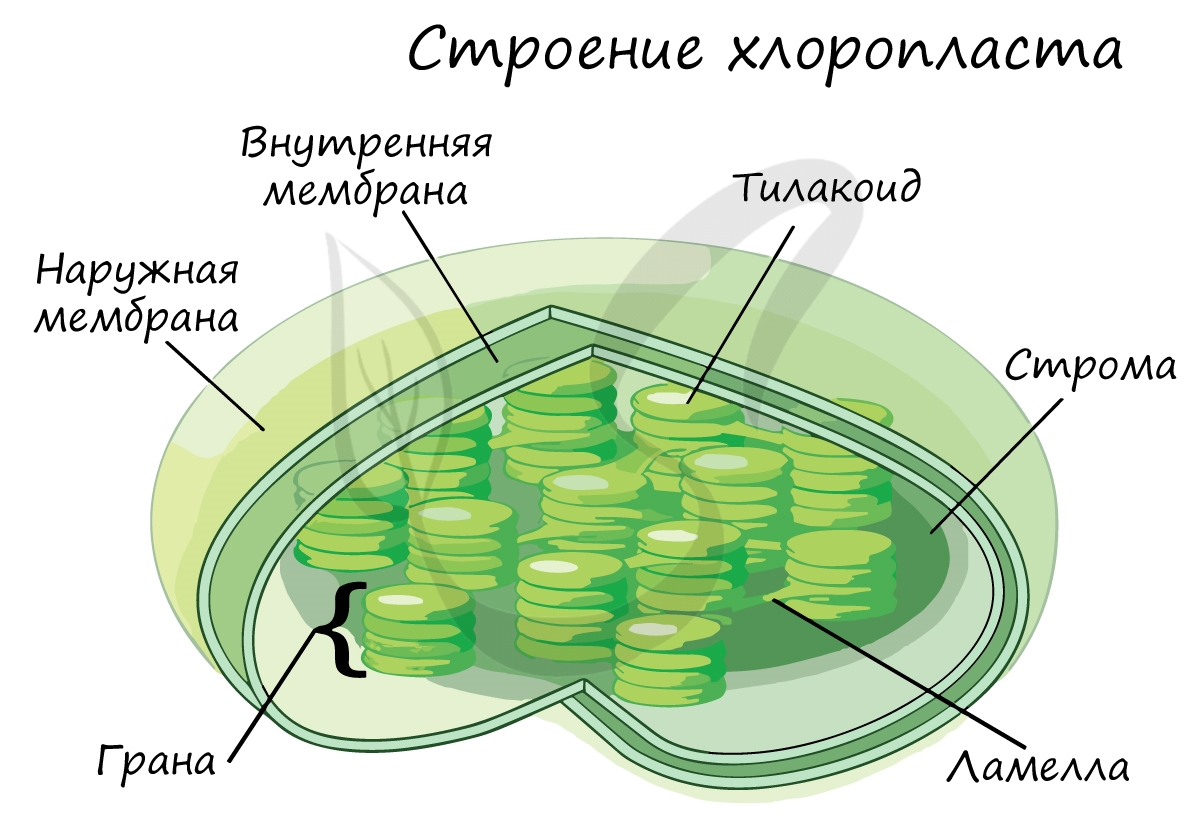
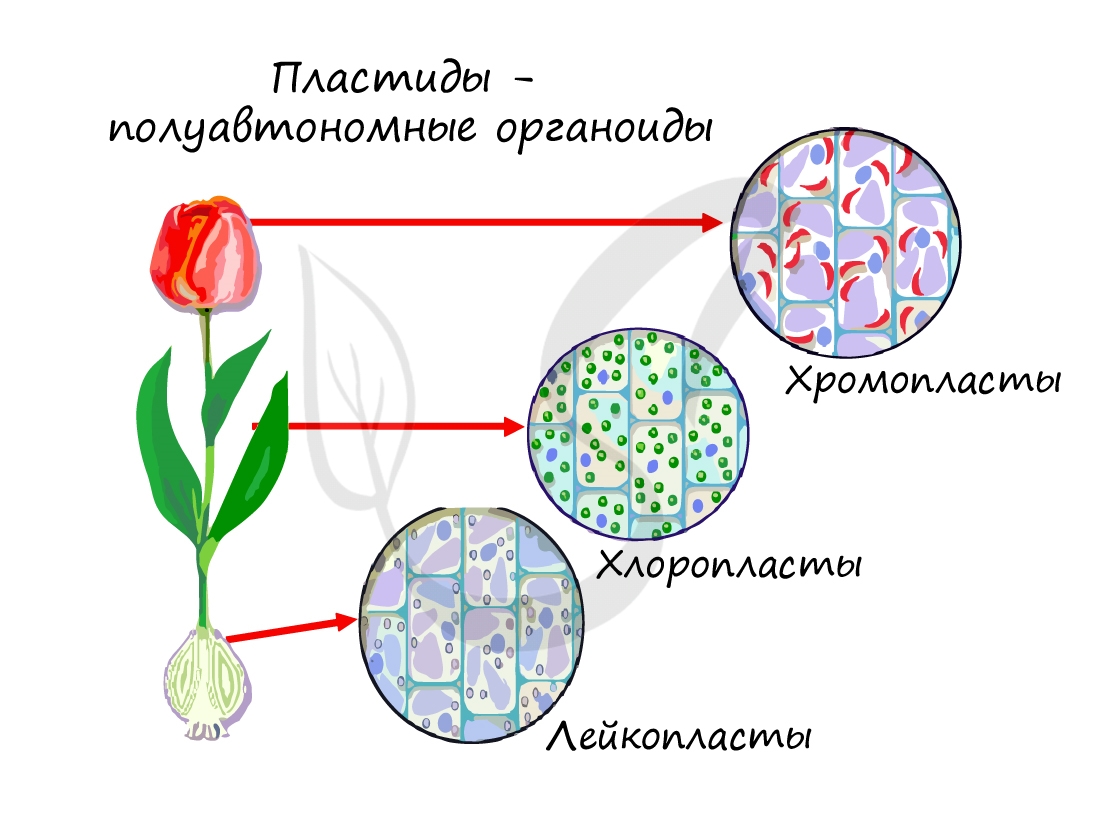
Так же, как и митохондрии, пластиды относятся к полуавтономным органоидам: в них имеется кольцевидная ДНК (находится в нуклеоиде), рибосомы.
Пластиды, которые содержат пигменты каратиноиды в различных сочетаниях. Сочетание пигментов обуславливает красную, оранжевую или желтую окраску. Находятся в плодах, листьях, лепестках цветков.
Хромопласты могут развиваться из хлоропластов: во время созревания плодов хлоропласты теряют хлорофилл и крахмал, в них активируется биосинтез каротиноидов.
Не содержат пигментов, образуются в запасающих частях растения (клубни, корневища). В лейкопластах накапливается крахмал, липиды (жиры), пептиды (белки). На свету лейкопласты могут превращаться в хлоропласты и запускать процесс фотосинтеза.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Гликолиз и глюконеогенез. Аэробное окисление глюкозы.
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
ГЛИКОЛИЗ. ГЛЮКОНЕОГЕНЕЗ. АЭРОБНОЕ ОКИСЛЕНИЕ ГЛЮКОЗЫ.
ГЛИКОЛИЗ
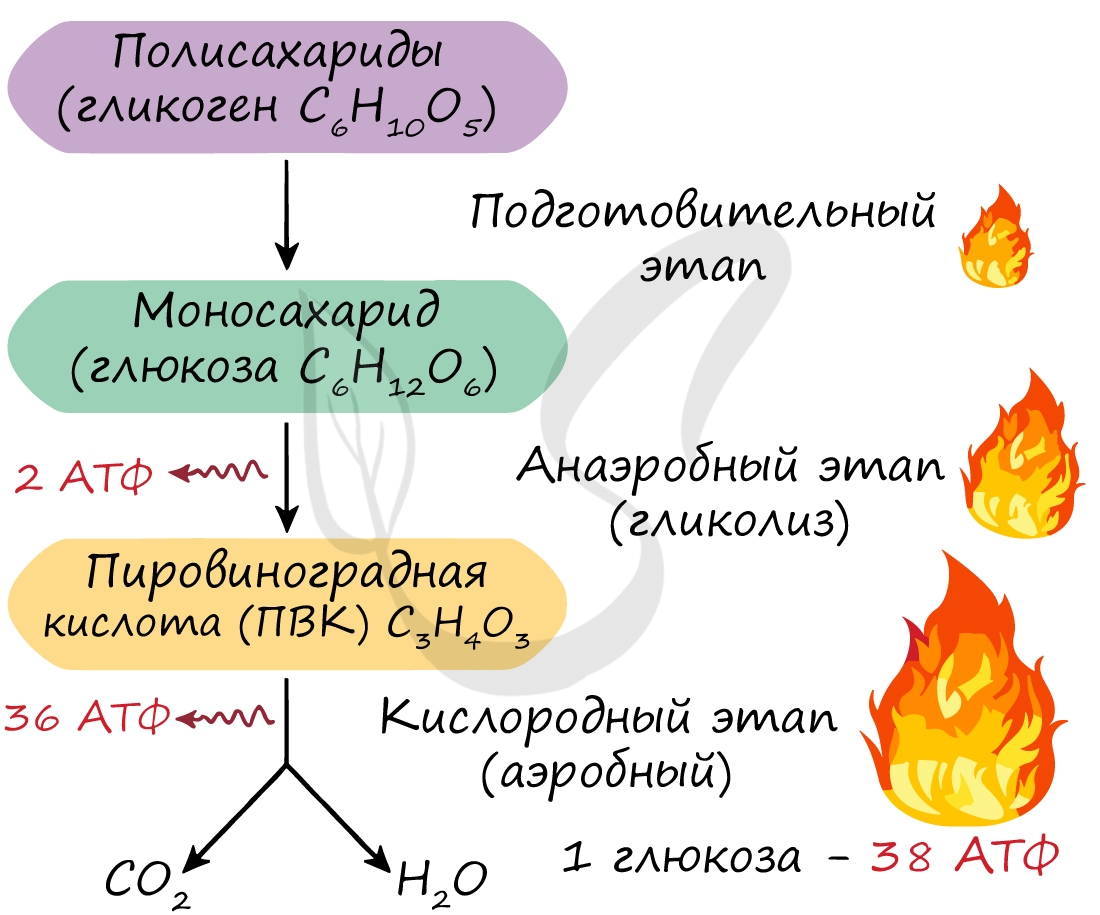
Гликолиз — это сложный ферментативный процесс расщепления глюкозы до двух молекул пирувата (аэробный гликолиз) или двух молекул лактата (анаэробный гликолиз, протекающий без потребления кислорода).
Суммарное уравнение анаэробного гликолиза:
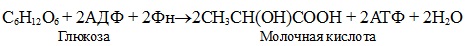
Гликолиз функционирует во всех живых клетках. Все ферменты локализованы в цитозоле, формируя полиферментный комплекс.
Гликолиз осуществляется в два этапа.
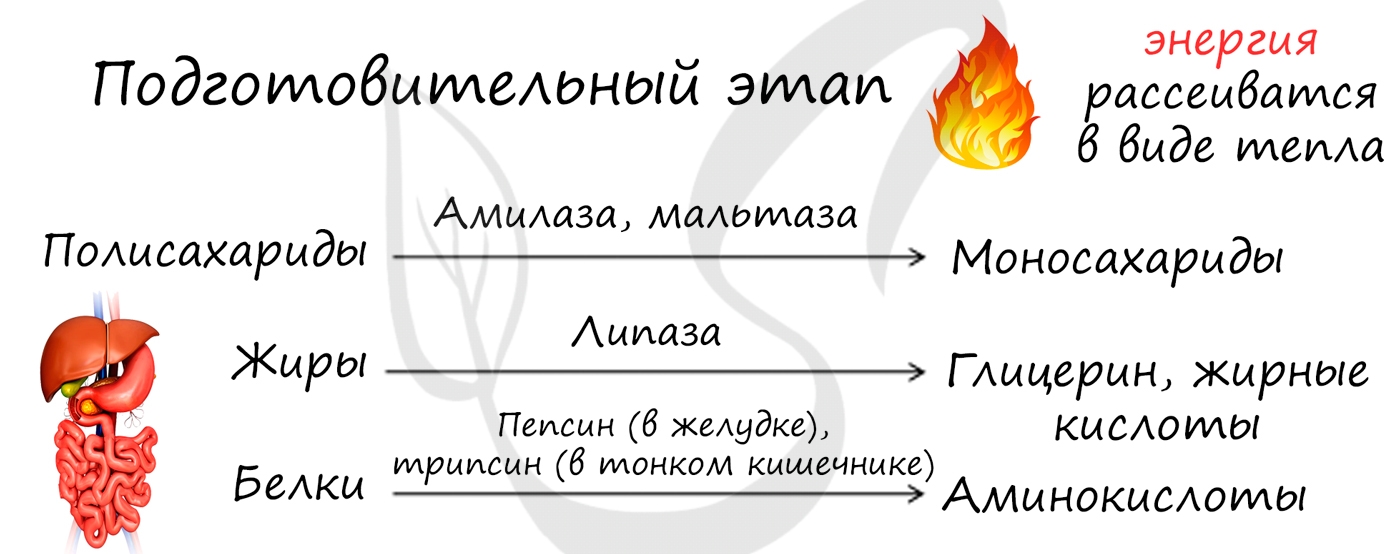
I. Подготовительный этап — дихотомический распад глюкозы на две молекулы глицеральдегид-3-фосфата. Превращения сопровождаются затратой 2 АТФ.
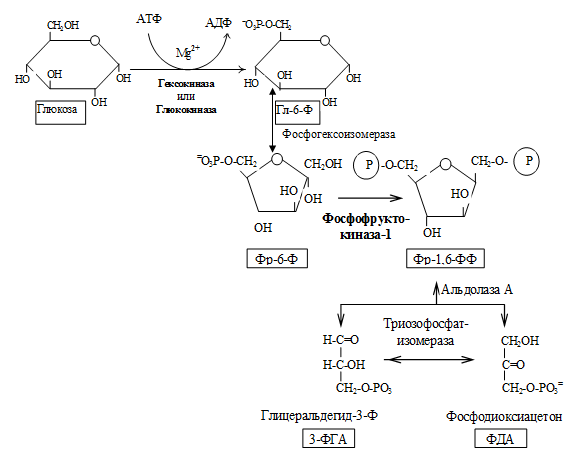
II. Этап гликолитической оксидоредукции — превращение глицеральдегид-3-фосфата в лактат. Включает окислительно-восстановительные реакции и реакции фосфорилирования, сопровождающиеся генерацией АТФ.
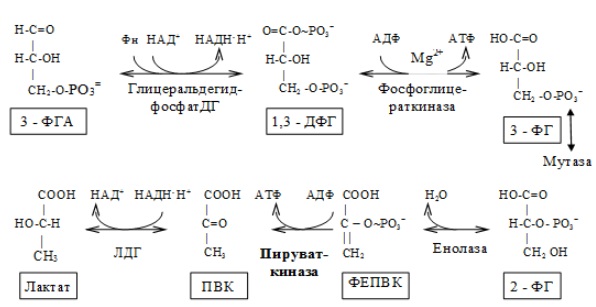
На втором этапе окисляются две молекулы глицеральдегид-3-фосфата, поэтому в реакциях впереди формулы субстрата следует ставить коэффициент 2.
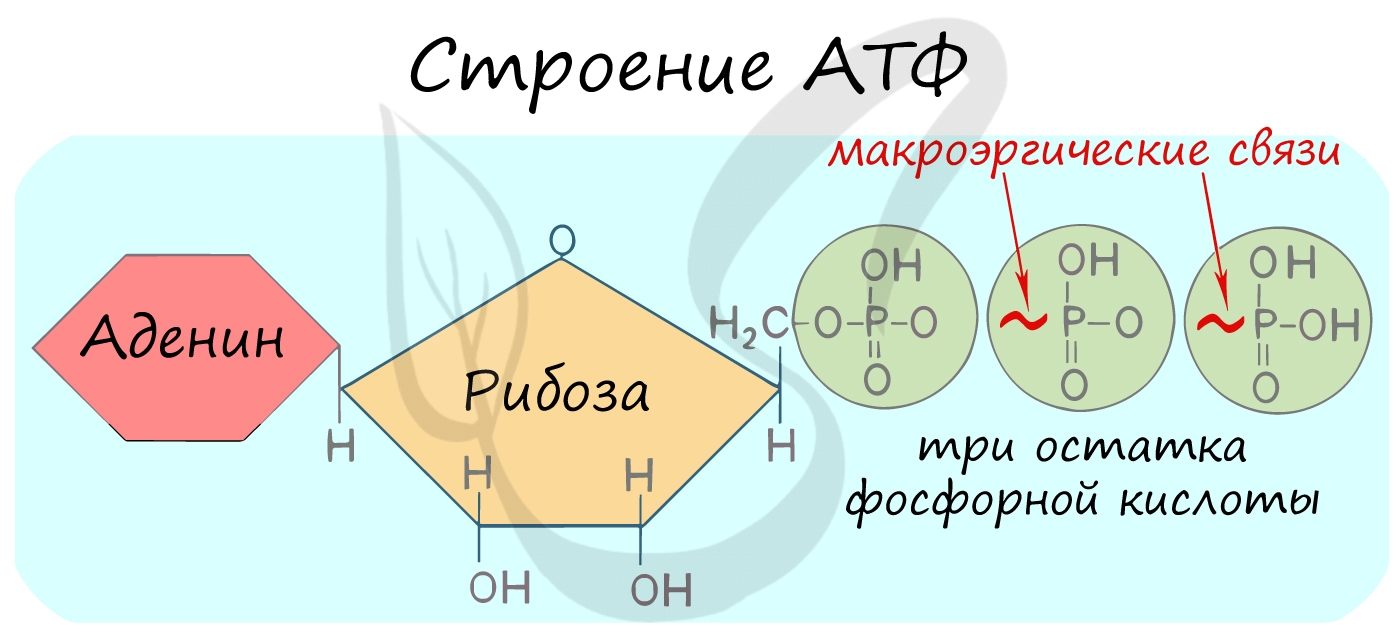
Энергетический баланс гликолиза — две молекулы АТФ на одну молекулу глюкозы. На I этапе гликолиза расходуются две молекулы АТФ для активирования субстрата (в гексокиназной и фосфофруктокиназной реакциях). На II этапе образуются четыре молекулы АТФ (в фосфоглицераткиназной и пируваткиназной реакциях). Синтез АТФ осуществляется путем субстратного фосфорилирования.
Ключевые ферменты гликолиза:
1. Гексокиназа — это регуляторный фермент гликолиза во внепеченочных клетках. Гексокиназа аллостерически ингибируется глюкозо-6-фосфатом. Глюкокиназа — регуляторный фермент гликолиза в гепатоцитах. Синтез глюкокиназы индуцируется инсулином.
2. Фосфофруктокиназа-1. Это главный ключевой фермент, катализирует реакцию, лимитирующую скорость всего процесса (наиболее медленная реакция). Синтез фермента индуцируется инсулином. Аллостерические активаторы — АМФ, АДФ, фруктозо-2,6-дифосфат. Уровень фруктозо-2,6-дифосфата увеличивается под действием инсулина и понижается под действием глюкагона. Аллостерические ингибиторы — АТФ, цитрат.
3. Пируваткиназа. Фермент активен в нефосфорилированной форме. Глюкагон (в гепатоцитах) и адреналин (в миоцитах) стимулируют фосфорилирование фермента, а значит инактивируют фермент. Инсулин, наоборот, стимулирует дефосфорилирование фермента, а значит активирует фермент. Аллостерический активатор — Фр-1,6-ФФ. Аллостерический ингибитор — АТФ, ацетилКоА. Синтез фермента индуцирует инсулин.
Биологическая роль гликолиза:
1. Генерирование АТФ. Гликолиз — единственный процесс в клетках, продуцирующий АТФ без потребления кислорода. Клетки, имеющие мало или не имеющие вообще митохондрий, получают АТФ только в ходе гликолиза.
Значение гликолиза для эритроцитов. Гликолиз — единственный процесс, продуцирующий АТФ в эритроцитах и поддерживающий их целостность и функции.
Наследственный дефект пируваткиназы сопровождается гемолитической анемией. При этой патологии эритроциты имеют от 5 до 25 % нормальной пируваткиназной активности и, следовательно, скорость гликолиза низкая.
Промежуточный продукт гликолиза в эритроцитах — 2,3-дифосфоглицерат (2,3-ДФГ) — понижает сродство гемоглобина к кислороду, способствуя диссоциации кислорода из оксигемоглобина и переходу его в ткани. Нарушения гликолиза в эритроцитах могут оказывать влияние на транспорт кислорода. Так, при недостаточности гексокиназы наблюдается понижение уровня 2,3-ДФГ и ненормально высокое сродство гемоглобина к кислороду. И наоборот, при недостаточности пируваткиназы содержание 2,3-ДФГ вдвое превышает норму, что обусловливает низкое сродство гемоглобина к кислороду.
2. Является источником углеводородных радикалов для процессов биосинтеза в клетках:
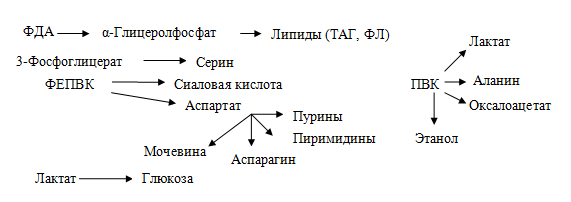
Патогенетическая взаимосвязь углеводов пищи и кариеса
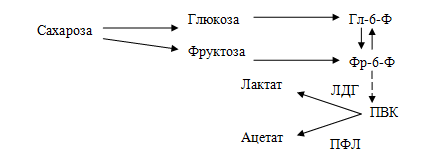
Потребление легкоферментируемых углеводов, в частности сахарозы, инициирует кислотную деминерализацию эмали зубов. У бактерий имеются два альтернативных пути использования пирувата: первый — путь восстановления ПВК в молочную кислоту с участием лактатдегидрогеназы (ЛДГ), второй — расщепление ПВК на уксусную и муравьиную кислоту с участием пируватформиатлиазы (ПФЛ).
Схема расщепления сахарозы под влиянием ферментов бактерий полости рта:
Зависимость выхода Са 2+ из зубов от рН ротовой жидкости:
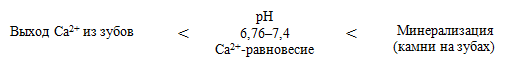
Аэробное окисление глюкозы
Это основной путь катаболизма глюкозы у аэробных организмов. Процесс осуществляется в три этапа. В аэробных условиях глюкоза окисляется до СО2 и Н2О.
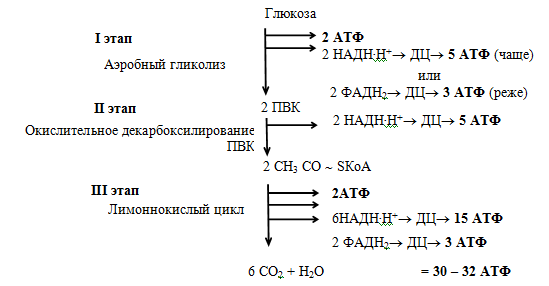
Энергетический баланс. Энергетический баланс аэробного окисления глюкозы —
30–32 моля АТФ на молекулу глюкозы.
ГЛЮКОНЕОГЕНЕЗ
Глюконеогенез — синтез глюкозы из соединений неуглеводной природы.
В организме взрослого человека за сутки может синтезироваться до 250 г глюкозы.
Глюконеогенез осуществляется главным образом в печени (синтезируетя до 90 % всей глюкозы), в корковом веществе почек и в энтероцитах (совсем незначительно).
Глюконеогенез стимулируется при длительном голодании, при ограничении поступления углеводов с пищей, в период восстановления после мышечной нагрузки, у новорождённых в первые часы после рождения.
Субстраты глюконеогенеза. Истинными субстратами глюконеогенеза являютя пируват, оксалоацетат, фосфодиоксиацетон, которые непосредственно включаются в этот процесс. Все вещества неуглеводной природы, дающие эти метаболиты, являются субстратами глюконеогенеза: лактат→ПВК, метаболиты цикла Кребса→ЩУК, глицерол→фосфодиоксиацетон, пропионил-КоА→метаболиты цикла Кребса→ЩУК, глюкогенные аминокислоты→ПВК или ЩУК. Главный источник субстратов глюконеогенеза — глюкогенные аминокислоты. К глюкогенным аминокислотам относятся все протеиногенные аминокислоты, кроме лейцина и лизина.
2ПВК + 4АТФ + 2ГТФ + 2НАДН . Н + + 2Н + 6Н2О → Глюкоза + 4АДФ + 2ГДФ + 6Фн + 2НАД +
Глюконеогенез протекает, в основном, по тому же пути, что и гликолиз, но в обратном направлении. Для обхода трех ключевых реакций гликолиза используются четыре специфических фермента глюконеогенеза.
Ключевые ферменты и ключевые реакции глюконеогенеза:
1. Пируваткарбоксилаза

2. Фосфоенолпируваткарбоксикиназа
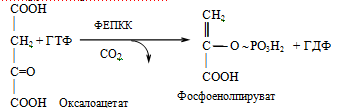
3. Фруктозо-1,6-бисфосфатаза
Фруктозо-1,6-бисфосфат + Н2О Фруктозо-6-фосфат + ФН
4. Глюкозо-6-фосфатаза
Глюкозо-6-фосфат + Н2О Глюкоза + ФН
Энергетический баланс. На синтез молекулы глюкозы из двух молекул пирувата расходуется 4АТФ и 2ГТФ (6АТФ). Энергию для глюконеогенеза поставляет процесс β-окис-ления жирных кислот.
Регуляция глюконеогенеза.
Глюконеогенез стимулируется в условиях гипогликемии при низком уровне инсулина и преобладании его антагонистов (глюкагона, катехоламинов, глюкокортикоидов).
1. Регуляция активности ключевых ферментов:
КоА (аллостерический активатор).
2. Регуляция количества ключевых ферментов: глюкокортикоиды и глюкагон индуцируют синтез ключевых ферментов, а инсулин — репрессирует.
3. Регуляция количества субстрата: количество субстратов глюконеогенеза увеличивается под действием глюкокортикоидов (катаболическое действие на белки мышечной и лимфоидной ткани, на жировую ткань), а также глюкагона (катаболическое действие на жировую ткань).
Биологическая роль глюконеогенеза:
1. Поддержание уровня глюкозы в крови. При длительном голодании (голодание более суток) глюконеогенез является единственным процессом, поставляющим глюкозу в кровь.
2. Возвращение лактата в метаболический фонд углеводов. Лактат, образующийся в процессе анаэробного окисления глюкозы в эритроцитах и скелетных мышцах, транспортируется кровью в печень и превращается в гепатоцитах в глюкозу. Это так называемый межорганный цикл Кори.
3. Предотвращение лактатного ацидоза, то есть в ходе глюконеогенеза лактат крови превращается в глюкозу.
Какой органоид является местом окисления глюкозы
Установите соответствие между характеристикой и органоидом клетки, к которому её относят. К каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
| ХАРАКТЕРИСТИКА | ОРГАНОИД КЛЕТКИ |