Органы дыхания земноводных
Условия обитания в воде и на суше очень различны, и организация амфибий глубочайшим образом отличается от организации их водных предков.
Органы дыхания. Влажность, постоянная для водной среды, на суше резко изменяется, и на большей части земной поверхности она относительно мала. Поскольку газообмен между организмом и средой возможен только через водную пленку, жабры, легко высыхающие на. воздухе, оказываются непригодными как органы дыхания для наземных животных. Они заменяются у амфибий легкими. В отличие от жабр легкие расположены внутри тела и защищены от высыхания.
 Рис. 2. AiexaminM акта дыхании лягушки: А Первый этап вдоха; Б — второй этап вдоха (стрелками указано направление движения воздуха); i — ротовая полость; 2 — язык; 3 — наружные ноздри; 4 — обонятельный мешок; 5 — хоаны; 6 — межчелюстная кость; 7 — вход в пищевод; 8 — легкое.
Рис. 2. AiexaminM акта дыхании лягушки: А Первый этап вдоха; Б — второй этап вдоха (стрелками указано направление движения воздуха); i — ротовая полость; 2 — язык; 3 — наружные ноздри; 4 — обонятельный мешок; 5 — хоаны; 6 — межчелюстная кость; 7 — вход в пищевод; 8 — легкое.
Легкие земноводных представляют собой парные мешки, полые внутри. Их тонкие стенки на внутренней поверхности имеют более или менее развитое ячеистое строение. Однако поверхность легких у земноводных еще невелика. Так, у большинства лягушек отношение ее к поверхности кожи равно 2:3, в то время как у млеко-питающих поверхность легкого в 50—100 раз больше поверхности кожи. Незначительное развитие легких у земноводных характеризует их как примитивных обитателей суши.
Дыхательные пути у амфибий развиты также слабо. У хвостатых они представлены в виде довольно длинной трубки — трахеи; у бесхвостых это лишь короткая трахейно-гортанная камера, непосредственно переходящая в полость легких.
Механизм дыхания у земноводных еще примитивного нагнетательного типа. Животное набирает воздух в ротовую полость, для чего открывает ноздри и опускает дно ротовой полости. Затем ноздри закрываются клапанами, дно ротовой полости поднимается и воздух нагнетается в легкие. Удаление воздуха из легких происходит благодаря действию брюшной мускулатуры спадению стенок легких. Механизм дыхания и отличает земноводных не только от рыб, но и от наземных позвоночных, дыхание которых осуществляется при помощи изменения объема грудной клетки.
Поскольку у земноводных относительная поверхность легких мала и они слабо вентилируются, то насыщение крови кислородом происходит не только через легкие, но и через кожу. Так, и пример, травяная лягушка получает через кожу 33% кислорода, а прудовая — 51%. Еще сложнее обстоит дело с удалением из организма углекислого газа. Благодаря недостаточной вентиляции легких углекислый газ скапливается в них затрудняет дальнейшую диффузию его из крови. Основная масса углекислого газа диффундирует в окружающую среду через кожу (у прудовой лягушки 86%). У высших наземных позвоночних, кожное дыхание ничтожно мало.
Покровы земноводных, функционирующие как дополнительный орган дыхания, отличаются в связи с этим рядом особенностей. Кожа у земноводных голая, что способствует свободному газообмену в кровеносных сосудах, образующих густую сеть. Так как обмен газам между организмом и средой идет только через водную пленку, которая у земноводных постоянно увлажняется слизью, вырабатываемой многочисленными железами. Кроме слизистых, в коже земноводных помещаются еще и ядовитые железы, секрет которых защищает животное от врагов и различных микроорганизмов.
Жизнь животных. В 7-ми т. / Гл. ред. В. Е. Соколов. Т. 5. Земноводные. Пресмыкающиеся / А. Г. Банников, И. С. Даревский, М. Н. Денисова и др.; Под ред. А. Г. Банникова. —2-е изд., перераб.—М.: Просвещение, 1985.—399с., ил., 32 л. ил.
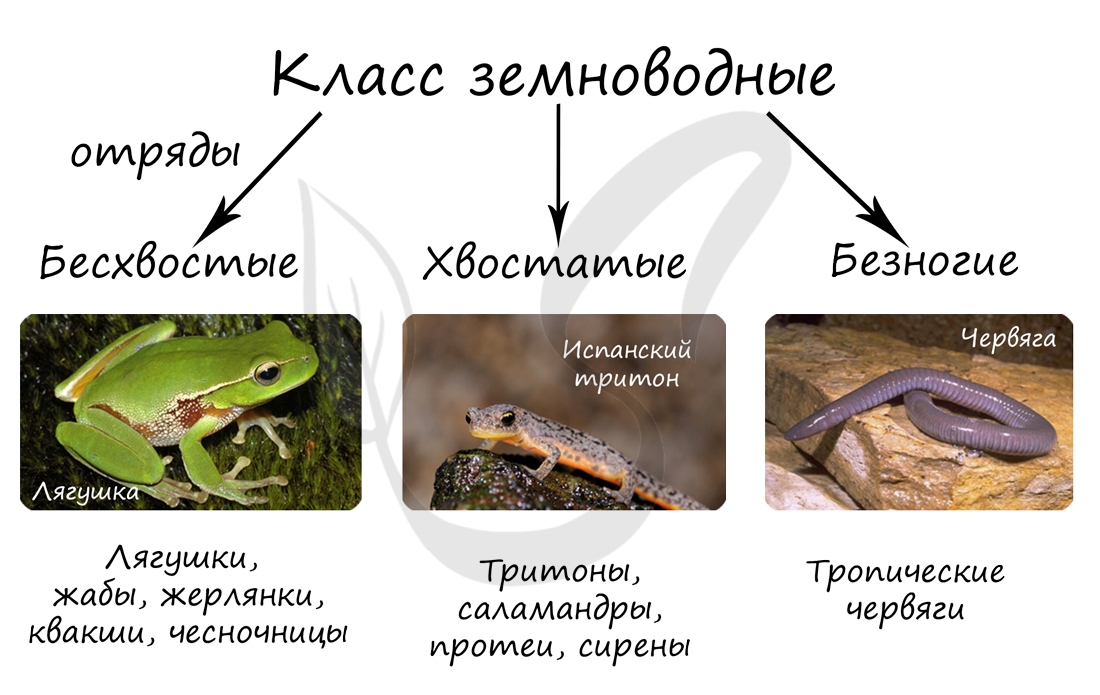
Класс земноводные
Для успешного изучения земноводных предлагаю начать с их классификации. Помните, что классификации это именно то, что раскладывает знания в голове «по полочкам», относитесь к ним с должным вниманием.
Давайте перечислим ароморфозы, которыми сопровождалось появление земноводных.
Ароморфозы земноводных
У ранее обсуждаемой нами рыбы латимерии (целаканта) плавники напоминали конечность пятипалого типа. У земноводных они преобразовываются в скелет передней и скелет задней конечностей, которые мы обсудим в данной статье подробно.

Мышцы дифференцируются (делятся) на мышечные пучки, становятся возможны более сложные и точные движения.

Легкие земноводных представлены в виде тонкостенных мешков, относительно небольшого объема.
Благодаря появлению шейного отдела становится возможным движение головой вверх-вниз, что было невозможным для рыб. Однако, подвижность головы значительно ограничена, и поворот головы из стороны в сторону невозможен.

Лягушка озерная

Покров двуслойный, состоит из эпидермиса и дермы. Эпидермис снабжен густой капиллярной сетью, благодаря которой у лягушки возможен газообмен через кожу. Кожа голая, содержит большое количество желез: она должна быть постоянно увлажнена для интенсивного газообмена.
Около 65% всего кислорода поступает через увлажненную кожу. Увлажнение кожи крайне важно, рекомендую запомнить следующий факт: при пересыхании кожи лягушки газообмен в ней прекращается, и лягушка погибает от недостатка кислорода (гипоксии).


Тело состоит из головы, туловища и двух пар конечностей. Передние конечности короче и слабее задних. Задние конечности служат для плавания, они удлинены и имеют плавательные перепонки между пальцами. Хорошо развита мышечная система: мышцы дифференцированы на отдельные пучки. В теле амфибий насчитывается до 350 мышц.
Скелет лягушки состоит из скелета головы, туловища, поясов конечностей и свободных конечностей. Скелеты поясов конечностей малоподвижны, в отличие от скелета свободной конечности, построенного по типу рычагов.
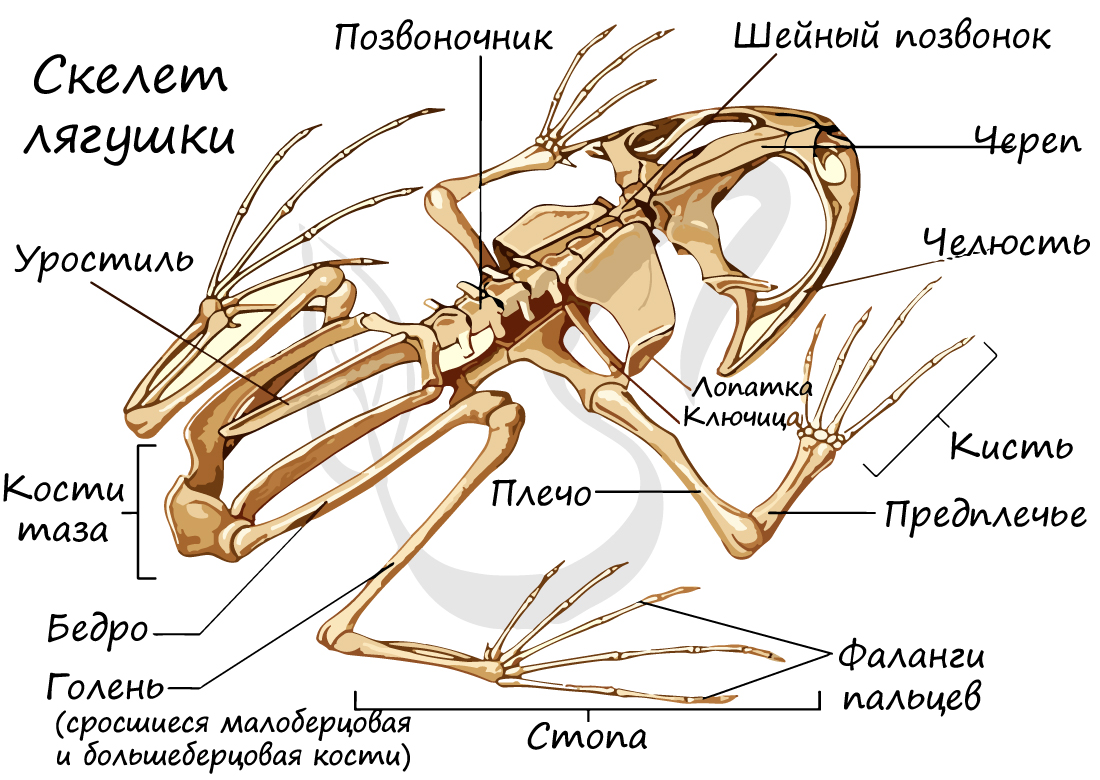
Имеются туловищный (7 позвонков) и хвостовой отделы позвоночника, доставшиеся земноводным еще от рыб. Однако появляются и новые отделы: крестцовый и шейный, оба содержащие по одному позвонку. Благодаря возникновению шейного отдела, у земноводных становится возможным поворот головы, что было невозможным для рыб.
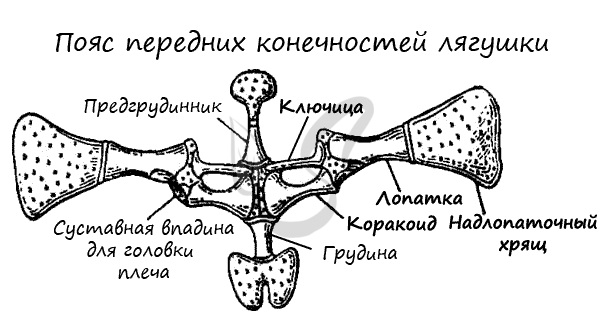
Скелеты поясов конечностей служат опорой для конечностей. В состав плечевого (переднего) пояса входят: лопатки, ключицы, вороньи кости (коракоиды), хрящи. Ребер нет, вследствие чего грудная клетка отсутствует, передний пояс лежит свободно в толще мышц, грудина не соединена при помощи ребер с позвоночником.
Пояс задних конечностей (тазовый) образован: подвздошными и седалищными костями, лобковым хрящом.
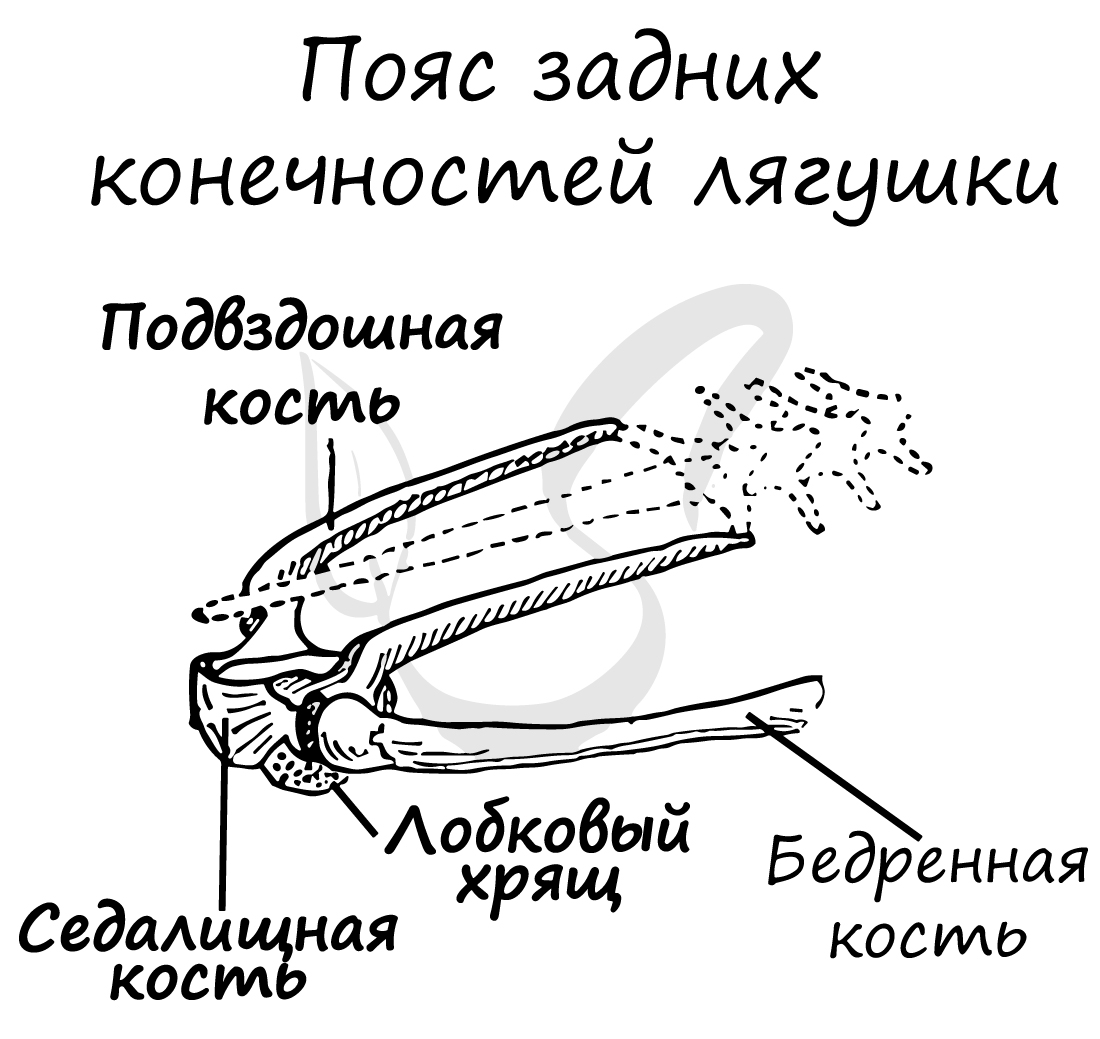
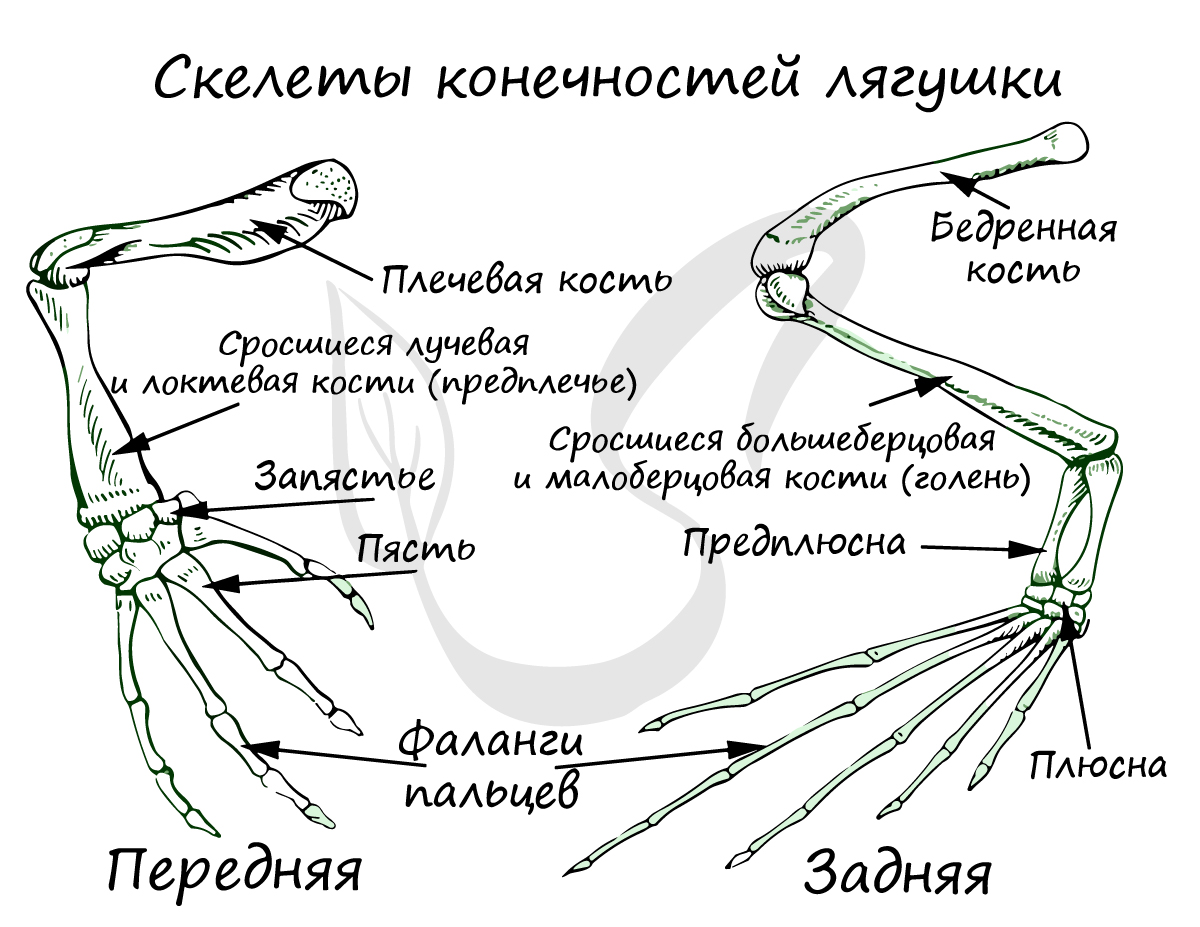
Скелет передней свободной конечности состоит из: плечевой кости, предплечья (образовано сросшимися лучевой и локтевой костями) и кисти (состоящей из запястья, пястья и фаланг пальцев).
Скелет задней свободной конечности образован бедром, голенью (состоит из сросшихся малоберцовой и большеберцовой костей), стопы (включает предплюсну, плюсну и фаланги пальцев).

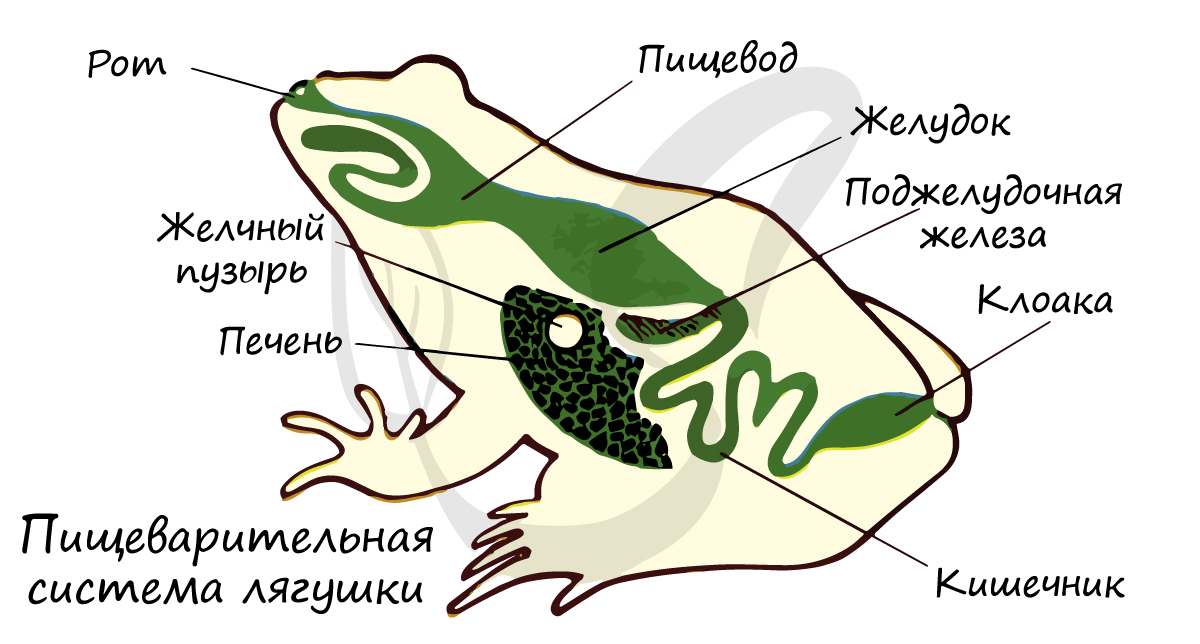
Короткий пищевод переходит в желудок, продолжающийся в тонкий кишечник, куда открываются протоки желчного пузыря, а также пищеварительных желез: печени и поджелудочной. Тонкая кишка переходит в толстую кишку, оканчивающуюся клоакой.
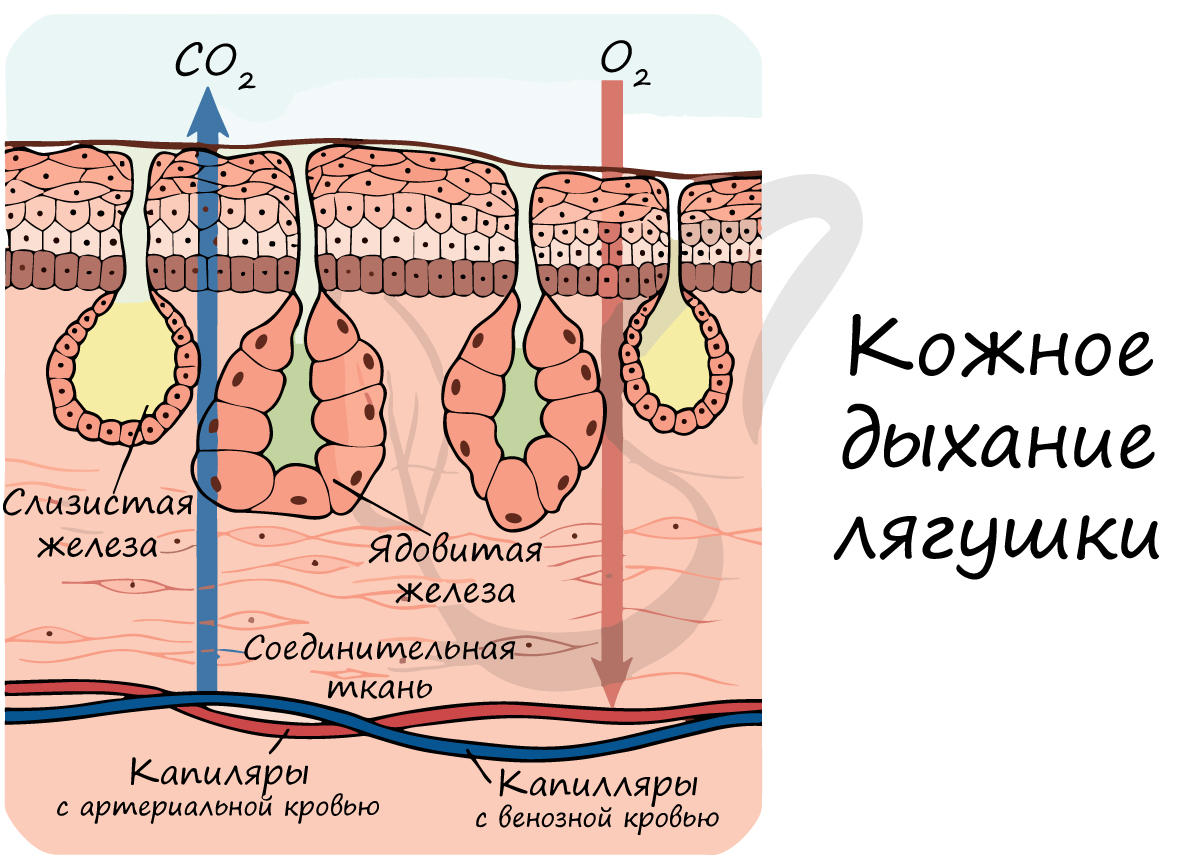
Легкие представленные тонкостенными мешками, стенка которых густо оплетена капиллярами, в которых происходит газообмен. Дыхательная поверхность легких невелика, они не способны полностью удовлетворить нуждам организма в кислороде, поэтому жизнь лягушки невозможна без кожного дыхания.
Воздух поступает в легкие и удаляется из них благодаря сокращениям мышц дна ротовой полости.

Осуществляется только при увлажненной коже, на поверхности которой растворяется кислород, после чего он проникает в капилляры кожи. Кожа помогает дышать лягушкам при погружении в воду: кислород поступает в кровь напрямую из воды.
При пересыхании кожи животное погибает, так как слабо развитые легкие не в состоянии обеспечить потребности организма в кислороде.
Также дыхание осуществляется эпителием, покрывающим ротовую полость, но это вносит незначительный вклад в процесс дыхания.
Изменения в кровеносной системе земноводных, по сравнению с рыбами, обусловлены заменой жаберного дыхания на легочное. Сердце имеет 3 камеры: 2 предсердия и 1 желудочек, сердце более крупное. Возникает второй (легочный) круг кровообращения.
Физиологически и анатомически сокращение сердца происходят таким образом, что более насыщенная кислородом артериальная кровь изгоняется из него в последнюю очередь и попадает в сонные артерии, идущие к головному мозгу. Таким образом, к головному мозгу направляется более насыщенная кислородом кровь, чем к остальным органам.
По мере образования в почке мочи, она поступает в мочеточники, ведущие в клоаку. Попавшая в клоаку моча стекает в мочевой пузырь, через стенки которого осуществляется всасывание воды. При сокращении стенок мочевого пузыря концентрированная моча выводится в клоаку, откуда выбрасывается во внешнюю среду.
Головной мозг земноводных отличается от мозга рыб полностью разделенным на два полушария и более развитым передним мозгом.
Мозжечок и средний мозг развиты гораздо слабее, так как земноводные малоподвижны и их движения относительно простые: они передвигаются по плоскости в «2D измерении». Перемещение рыб в таком случае можно обозначить как в «3D измерении». Это упрощенное, но вполне справедливое сравнение.
Свободные конечности земноводных позволяют им совершать более разнообразные движения в отличие от циклических поворотов плавников и тела у рыб.
Меняется форма хрусталика. Если у рыб он был шарообразный, то у земноводных его форма становится более выпуклой (как и форма роговицы). Хрусталик приобретает форму двояковыпуклой линзы. Амфибии видят дальше рыб.
Однако аккомодация, настройка глаза на наилучшее видение объекта, остается на том же уровне, что и у рыб: достигается только за счет перемещения хрусталика вперед и назад. Эти перемещения помогает совершать особая ресничная мышца, возникшая у земноводных.

Наземная среда меняет и характер передачи звуков: теперь он передается через колебания воздуха. Если рыбам в воде было достаточно внутреннего уха для улавливания колебаний воды, то земноводным этого мало. У них впервые возникает среднее ухо, которое снабжено барабанной перепонкой, чувствительной к колебаниям воздуха.

Лягушки раздельнополые животные, оплодотворение наружное. Размножение и начальные этапы развития земноводных происходят в воде, поэтому амфибий крайне редко можно встретить в засушливых местах, где водоемы отсутствуют.
Во время оплодотворения самец удерживает самку сзади и надавливает передними конечностями на ее брюшко, что приводит к выметыванию икры. Эта икра сразу же оплодотворяется семенной жидкостью самца.
Головастики


Значение земноводных
Земноводные, как и все живые организмы, являются звеном в цепи питания (консументами). Они уничтожают многих кровососущих насекомых, а также насекомых, которые наносят вред культурным растениям.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Амфибии Земноводные строение
Амфибии Земноводные животные класс сердце строение список система признаки особенности бесхвостое органы виды отличия кожа отделы
Латинское название Amphibia
 Амфибии Земноводные
Амфибии Земноводные
Общая характеристика
Амфибии или Земноводные — немногочисленная группа наиболее примитивных наземных позвоночных. Подавляющее большинство обитают, в зависимости от стадий жизненного цикла, то в воде, то на суше. В течение жизни амфибии, как правило, претерпевают метаморфоз, превращаясь из чисто водных личинок во взрослые формы, обитающие большей частью вне воды. В этой связи происходит смена жаберного дыхания на легочное, соответственно видоизменяется кровеносная система, появляются конечности типа пятипалой, существенно видоизменяется система органов чувств. Однако и у взрослых форм степень приспособления к жизни на суше в общем невелика. Легкие развиты слабо, и в качестве дополнительного органа дыхания функционирует еще и кожа. Трехкамерное сердце не обеспечивает полного разделения крови на артериальную и венозную, и в большинстве частей тела по артериям течет смешанная кровь. Конечности хотя и устроены по типу пятипалой, но развиты слабо и не могут удерживать тело в приподнятом над субстратом положении. Наконец, громадное большинство размножается в воде путем откладывания икры и оплодотворения ее вне тела матери (т. е. как и у рыб).
Общее число земноводных современных видов равно примерно 2100—2600; они объединяются в три отряда:
1. Хвостатые (Caudata, или Urodela).
3. Бесхвостые (Anura, или Ecaudata).
Кожные покровы
Кожа всех амфибий голая, лишенная наружного покрова из костных или роговых чешуй. Эпидермис очень богат железами, которые, в отличие от таковых рыб, многоклеточны. Значение кожных желез многообразно. Они обеспечивают наличие на поверхности кожи жидкостной пленки, без которой невозможен газообмен при кожном дыхании. Эта пленка в некоторой мере предохраняет тело от иссушения. Выделения некоторых кожных желез обладают бактерицидными свойствами и защищают от проникновения через кожу патогенных микробов. Ядовитые кожные железы в некоторой мере защищают от хищников. Установлено, что, вопреки прежнему мнению, верхний слой эпидермиса у амфибий, особенно у бесхвостых, ороговевает (В. Е. Соколов, 1964; Spearman, 1968), Наиболее сильно это развито у жаб, у которых на спине роговой слой составляет примерно 60% всей толщи эпидермиса. У большинства наличие слаборазвитых ороговений эпидермиса не препятствует проникновению через кожу воды, и амфибии, находясь в водоеме, постоянно «пьют воду кожей».
Позвоночник у земноводных в связи с полуназемным образом жизни более расчленен, чем у рыб. Он состоит из шейного, туловищного, крестцового и хвостового отделов. Шейный отдел представлен одним позвонком, его тело невелико и несет две сочленовные ямки, при помощи которых позвонок сочленяется с черепом. Число туловищных позвонков различно. Наименьшее число их у бесхвостых (обычно 7), наибольшее — у безногих (более ста). Единственный крестцовый позвонок (у безногих отсуствующий) несет длинные поперечные отростки, к которым причленяются подвздошные кости таза. Хвостовой отдел наиболее типично выражен у хвостатых, у безногих он очень мал, а у бесхвостых представлен косточкой — уростилем, которая во время эмбрионального развития закладывается в виде ряда отдельных позвонков.
Форма позвонков у низших амфибий (безногие, низшие хвостатые) бывает амфицельной; в этом случае пожизненно сохраняется хорда. У бесхвостых позвонки процельные, т.е. вогнутые спереди и выгнутые сзади, у высших хвостатых — опистоцельные, т.е. выгнутые спереди и вогнутые сзади. Только у крайне примитивной новозеландской лягушки лиопелъмы (Liopelma) позвонки амфицельные. Настоящие ребра, но очень короткие есть только у безногих; у хвостатых развиваются короткие «верхние» ребра, у бесхвостых ребер нет вовсе.
Мозговая коробка земноводных в значительной своей части пожизненно остается хрящевой. Это обусловливается слабым развитием хондральных окостенений и накладных костей. В первичной мозговой коробке развиваются следующие хондральные кости. В затылочной области только две боковые затылочные кости (exoccipitale); места, соответствующие основной и верхней затылочным костям рыб, остаются хрящевыми. В области слуховой капсулы формируется одна небольшая ушная кость (prooticum), большая же часть капсулы остается хрящевой. В передней части глазницы у бесхвостых развивается клинообонятельная кость (sphenethmoideum); у хвостатых эта кость парная; обонятельная капсула сохраняет хрящевой характер.
Покровных костей, как сказано, также немного. Крышу черепа земноводных составляют теменные (parietale) и лобные (frontale) кости, которые у бесхвостых срастаются в лобно-теменные кости (frontoparietale). К переди от них находятся носовые кости (nasale), у безногих они срастаются с предчелюстными костями (praemaxillare). По бокам задней части черепа расположены чешуйчатые кости (squamosum), особенно сильно развитые у безногих. Большой парасфеноид (parasphenoideum) выстилает дно черепа, а вперед от него лежат парные сошниковые кости (vomer).
В формировании дна черепа принимают участие и кости висцерального скелета: нёбные (palatinum) и крыловидные (pterygoideum). Первая прилегает к сошнику, вторая — к чешуйчатой кости.
Череп амфибий аутостилический, т. е. нёбноквадратный хрящ непосредственно прирастает к мозговому черепу. Упомянутые выше нёбные и крыловидные кости развиваются на нижней поверхности нёбноквадратного хряща. Функции верхних челюстей выполняет, как и у костистых рыб, костная дуга, состоящая из предчелюстных или межчелюстных костей (praemaxillare или intermaxillare) и верхнечелюстных костей (maxillare). Эта костная дуга лежит несколько к наружи от дуги, образуемой нёбноквадратными хрящами.
Нижняя челюсть представлена меккелевым хрящом, который прикрыт снаружи зубной (dentale) и угловой (angulare) костями.
В связи с аутостилией черепа подъязычная дуга не принимает участия в прикреплении челюстного аппарата к черепу. Верхний элемент этой дуги— гиомандибуляре — превращен в маленькую кость — стремя (stapes), — которая, как и у рыб, верхним своим концом упирается в слуховую капсулу. В связи с формированием полости среднего уха эта кость находится внутри указанной полости и выполняет роль слуховой косточки.
Нижние элементы подъязычной дуги и жаберных дуг видоизменяются в подъязычную пластинку и ее рожки. Пластинка эта расположена между ветвями нижней челюсти. Передние ее рожки, загибаясь вверх и охватывая с боков кишечную трубку, прикреплены к слуховым капсулам.
Таким образом, видно, что череп амфибий отличается от черепа большинства костных рыб: 1) слабым развитием хондральных и кожных окостенений; 2) аутостилией; 3) видоизменением подъязычной и жаберных дуг, превращенных частью в слуховой, частью в подъязычный аппарат; 4) редукцией жаберной крышки. Следовательно, наиболее существенные изменения наблюдаются в области висцерального черепа, и они связаны с переходом животных к полуназемному существованию (утеря жаберного аппарата, появление первой слуховой косточки, развитие своеобразного подъязычного скелета).
Пояса конечностей
Плечевой пояс имеет вид дуги, обращенной вершиной к брюшной поверхности животного. Каждая половина дуги (левая и правая) состоит из следующих основных элементов. Верхняя (спинная) часть представлена лопаткой (scapula), заканчивающейся широким надлопаточным хрящом. Нижняя (брюшная) часть состоит из коракоидa (coracoideum) и лежащего впереди него прокоракоида (procoracoideum). Перечисленные три элемента пояса сходятся в точке прикрепления плечевой кости и фомируют сочленовную ямку. Впереди от места соединения левого и правого коракоидов находится предгрудинa (omosternum), а сзади — грудина (sternum). Обе эти кости заканчиваются хрящами. У бесхвостых между предгрудиной и лопаткой расположена тон¬кая палочковидная ключица (clavicula). В связи с отсутствием или неполным развитием ребер грудной клетки у амфибий нет, и плечевой пояс лежит свободно в толще мускулатуры.
Тазовый пояс образован тремя парными элементами, сходящимися в области вертлужной впадины, которую они и образуют. Длинные подвздошные кости (ilium) своими проксимальными концами прикреплены к поперечным отросткам крестцового позвонка. Направленный вперед и вниз лобковый элемент пояса (pubis) у лягушек остается хрящевым. Сзади него расположена седалищная кость (ischium). Такое расположение элементов тазового пояса характерно для всех наземных позвоночных.
Скелет свободных конечностей
Скелет свободных конечностей типичен для наземных позвоночных и существенно отличен от скелета конечностей рыб. В то время как конечности рыб представляют в схеме простые одночленные рычаги, перемещающиеся только относительно корпуса тела, конечности наземных позвоночных представляют рычаги многочленные. В этом случае не только вся конечность перемещается относительно тела, но перемещаются относительно друг друга и отдельные элементы конечности.
Скелет конечностей хвостатых амфибий почти полностью соответствует приведенной схеме. У лягушек же наблюдаются некоторые отклонения, основными из которых будут следующие: оба элемента предплечья и голени срастаются в одну кость, срастается между собой и большая часть костей запястья и предплюсны, перед первым пальцем задней конечности имеется рудимент добавочного пальца (praehallux). Эти особенности имеют вторичный характер и связаны с приспособлением лягушек к передвижению прыжками.
Мышечная система
Мышечная система существенно отличается от мышечной системы рыб двумя основными особенностями, связанными с передвижением животных при помощи пятипалых конечностей и в известной мере по твердому субстрату. Во-первых, развивается мощная и сложно организованная мускулатура на свободных конечностях. (Вспомним, что практически у всех рыб мускулатура, приводящая в движение конечности, располагалась не на самых конечностях, а на корпусе тела.) Во-вторых, в связи с более сложными движениями мускулатура тела значительно более дифференцирована и характерная для рыб сегментация у амфибий нарушена. Метамерное расположение мышечных сегментов можно видеть у бесхвостых земноводных только в немногих местах туловища. У хвостатых земноводных и безногих метамерия мышечной системы выражена более четко.
Органы пищеварения
Ротовая щель земноводных ведет в обширную ротоглоточную полость, которая, сужаясь, переходит в пищевод. В ротоглоточную полость открываются хоаны, евстахиевы отверстия (полости среднего уха) и гортанная щель. Сюда же открываются протоки слюнных желез (отсутствующие у рыб), которые, однако, служат только для смачивания пищевого комка и секрет которых не воздействует на пищу химически. На дне ротоглоточной области расположен настоящий язык, имеющий собственную мускулатуру. Форма языка земноводных разнообразна. У некоторых хвостатых он прикреплен неподвижно, у других имеет вид гриба, сидящего на тонкой ножке. У лягушек язык прикреплен к дну рта своим передним концом, а свободная его часть в спокойном состоянии обращена внутрь. У всех язык выделяет клейкое вещество и служит для ловли мелких животных. Не имеют языка только немногие амфибии.
Зубы земноводных имеют вид мелких однообразных конусов, вершины которых несколько загнуты назад. Зубы сидят на межчелюстных, верхнечелюстных костях, на сошнике, а у некоторых и на нижней челюсти. Некоторые виды, например жабы, не имеют зубов на костях челюстей. При глотании проталкиванию пищевого комка из ротоглоточной области в пищевод помогают глазные яблоки, которые отделены от этой полости только тонкой слизистой оболочкой и которые с помощью специальных мышц могут несколько втягиваться внутрь ротоглотки.
Короткий пищевод земноводных впадает в сравнительно слабо отграниченный желудок. Собственно кишечник относительно более длинный, чем у рыб. В петле переднего (тонкого) отдела лежит поджелудочная железа (pancreas). Крупная печень имеет желчный пузырь, ее проток впадает в переднюю часть тонкой кишки (в так называемую двенадцатиперстную кишку). В желчный проток впадают и протоки поджелудочной железы, которая, таким образом, самостоятельного сообщения с кишечником не имеет.
Второй отдел кишечника — толстый — отграничен от тонкого отдела нечетко. Наоборот, третий — прямой — отдел хорошо обособлен. Он открывается в клоаку.
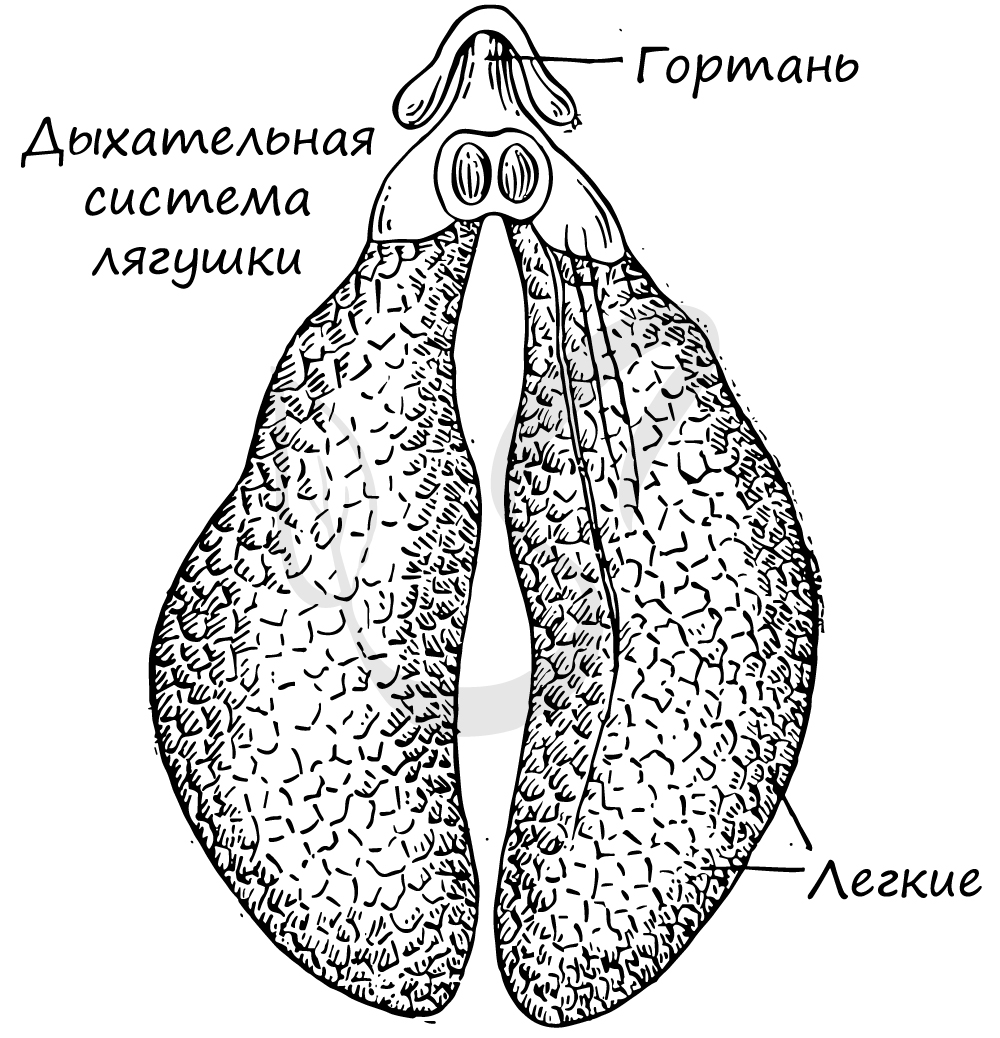
Органы дыхания
Органы дыхания земноводных разнообразны. Во взрослом состоянии большинство видов амфибий дышат легкими и через кожу. Легкие представляют парные мешки с тонкими ячеистыми стенками. В связи с несовершенством легких значение кожного дыхания очень велико. Отношение поверхности легких к поверхности кожи у амфибий равно 2 : 3 (в то время как у млекопитающих внутренняя поверхность легких в 50—100 раз больше поверхности кожи). У зеленой лягушки через кожу поступает 51 % кислорода, потребляемого для окисления крови, и 49% его поступает через легкие. Еще большее значение кожного дыхания видно при учете выделяемой организмом углекислоты: кожное дыхание — 86%, легочное дыхание — 14%.
Кожное дыхание имеет большое функциональное значение не только в связи с несовершенством легких, но и как приспособление, обеспечивающее окисление крови при длительном нахождении животного в воде, например во время спячки или затаивания в водоеме при преследовании наземными хищниками. В этих случаях осуществляется только кожное дыхание и правое предсердие (куда через полую вену вливается окисленная кровь кожной вены) становится артериальным, а левое — венозным.
У американских безлегочных саламандр и у нашего дальневосточного тритона легкие полностью атрофированы, и газообмен у них целиком про исходит через кожу и слизистую ротовой полости.
Личинки амфибий дышат при помощи ветвистых наружных жабр, которые у подавляющего большинства видов в последующем исчезают. Но у своеобразных протеев и сирен они сохраняются пожизненно. Амфиумы же во взрослом состоянии наряду с легкими имеют и внутренние жабры.
Механизм легочного дыхания земноводных в связи с отсутствием грудной клетки весьма своеобразен. Роль насоса выполняет ротоглоточная область, дно которой то опускается (воздух при открытых ноздрях засасывается), то поднимается (воздух при закрытых ноздрях проталкивается в легкие).
В этой связи череп амфибий при небольшой высоте чрезвычайно широк: эффективность легочного дыхания будет тем большей, чем шире расстояние между ветвями нижней челюсти.
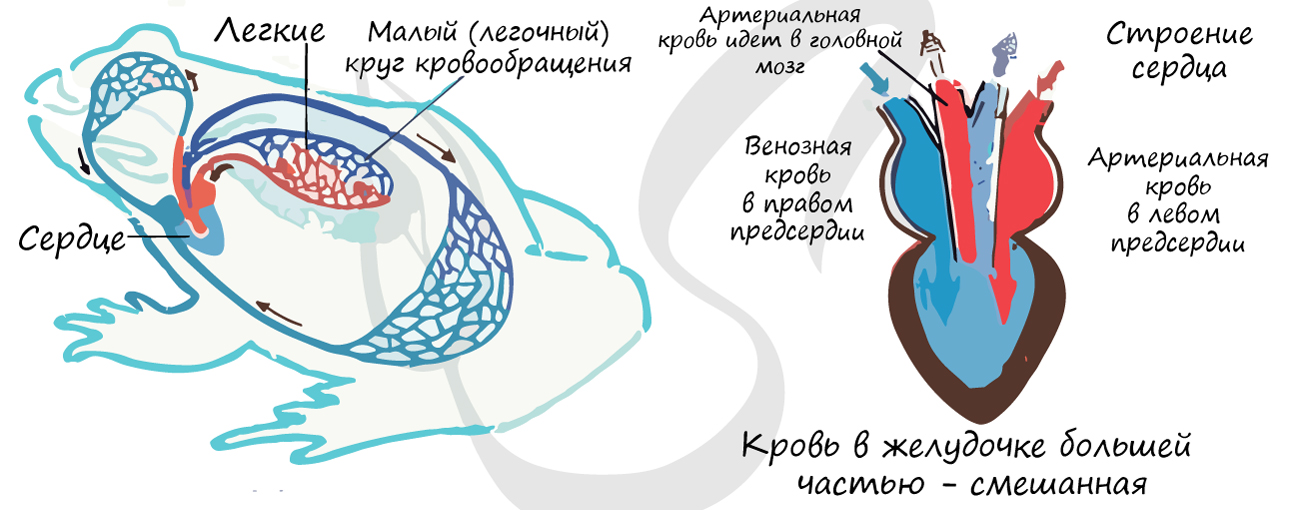
Кровеносная система
Сердце всех амфибий трехкамерное, состоит из двух предсердий и одного желудочка. У низших форм (безногие и хвостатые) левое и правое предсердия разделены не полностью. У бесхвостых перегородка между предсердиями полная, но у всех земноводных оба предсердия сообщаются с желудочком одним общим отверстием. Кроме указанных основных отделов сердца, имеется венозная пазуха, сообщающаяся с правым предсердием, и артериальный конус.
У высших бесхвостых амфибий от артериального конуса берут начало три пары артериальных дуг. Первая пара (считая от головы к хвосту), гомологичная первой паре жаберных артерий рыб, несет кровь к голове и представляет сонные артерии (arteria carotis). Она отходит от брюшной стороны артериального конуса. Вторая пара, также отходящая от брюшной стороны артериального конуса, гомологична второй паре жаберных сосудов рыб и носит название системных дуг аорты. От нее отходят подключичные артерии (arteria subclavia), несущие кровь к плечевому поясу и передним конечностям. Правая и левая системные дуги, описав полукруг, соединяются вместе и образуют спинную аорту (aorta dorsalis), расположенную под позвоночником и дающую начало веткам, идущим к внутренним органам. Последняя, третья, пара, гомологичная четвертой жаберной дуге рыб, отходит не от брюшной, а от спинной стороны артериального конуса. Она несет кровь в легкие и пред-ставляет легочные артерии (arteria pulmonalis). От каждой легочной артерии отходит по большой ветке, несущей венозную кровь в кожу для окисления. Это кожные артерии (arteria cutanea).
У хвостатых амфибий, у которых пожизненно сохраняются жабры, схема кровообращения очень близка к таковой рыб и личинок высших амфибий. От брюшной аорты амфибий отходят четыре пары дуг. Сонные артерии отходят от первой дуги (от выносящей ее части); с появлением же легких образуются легочные артерии, берущие начало от четвертой дуги. Как видно, схема кровообращения в этом случае почти тождественна таковой двоякодышащих рыб.
Венозная система низших амфибий похожа на венозную систему двоякодышащих рыб. Хвостовая вена делится на две воротные вены почек, из которых кровь поступает в непарную заднюю полую вену (vena cava posterior) и в парные задние кардинальные вены. Последние на уровне сердца сливаются с парными яремными, подключичными и кожными венами и образуют кювьеровы протоки, изливающие кровь в венозную пазуху. Полая вена принимает в себя печеночную вену (vena hepatica) и также впадает в венозную пазуху. От кишечника кровь собирается по подкишечной и брюшной венам, которые, Сливаясь, образуют воротную вену печени. Из печени кровь поступает по уже упомянутой печеночной вене.
У бесхвостых амфибий кардинальные вены не сохраняются и вся кровь из туловищной области собирается в конечном счете в заднюю полую вену, впадающую в венозную пазуху. Так же как и в предыдущем случае, имеются брюшная и подкишечная вены, образующие воротную систему кровообращения в печени. В связи с отсутствием кардинальных вен не образуются кювьеровы протоки. Яремные вены, сливаясь с подключичными, образуют в этом случае парную переднюю полую вену (vena cava anterior), впадающую в венозный синус (пазуху). В верхние полые вены впадают и кожные вены соответствующей стороны, которые несут не венозную, а артериальную кровь.
Легочные вены впадают непосредственно в левое предсердие.
Рассмотрим в заключение схему циркуляции крови земноводных.
Венозная кровь земноводных (с довольно значительной примесью окисленной крови, поступающей из кожных вен в передние полые вены) изливается в венозную пазуху (синус), а оттуда в правое предсердие. Одновременно в левое предсердие по легочным венам поступает чистая артериальная кровь. При сокращении предсердий венозная и артериальная кровь проталкивается через общее для обоих предсердий отверстие в желудочек. При сокращении желудочка в артериальный конус (в связи с его отхождением от правой части желудочка) поступает сначала более венозная кровь, идущая далее в открытое отверстие кожно-легочных артерий. Отверстия остальных артериальных дуг в это время закрыты спиральным клапаном артериального конуса. При дальнейшем сокращении желудочка давление в артериальном конусе возрастает, спиральный клапан сдвигается и открываются отверстия системных дуг, через которые поступает смешанная кровь из центральной части желудочка. Наиболее артериальная кровь из левой части желудочка, выходящая в артериальный конус, в последнюю очередь не попадает в легочные и системные дуги, которые уже заполнены кровью. Дальнейшее сдвигание спирального клапана освобождает устья сонных артерий, куда и проходит наиболее окисленная кровь. При всем этом полного разделения потоков артериальной и венозной крови все же нет.
Нервная система
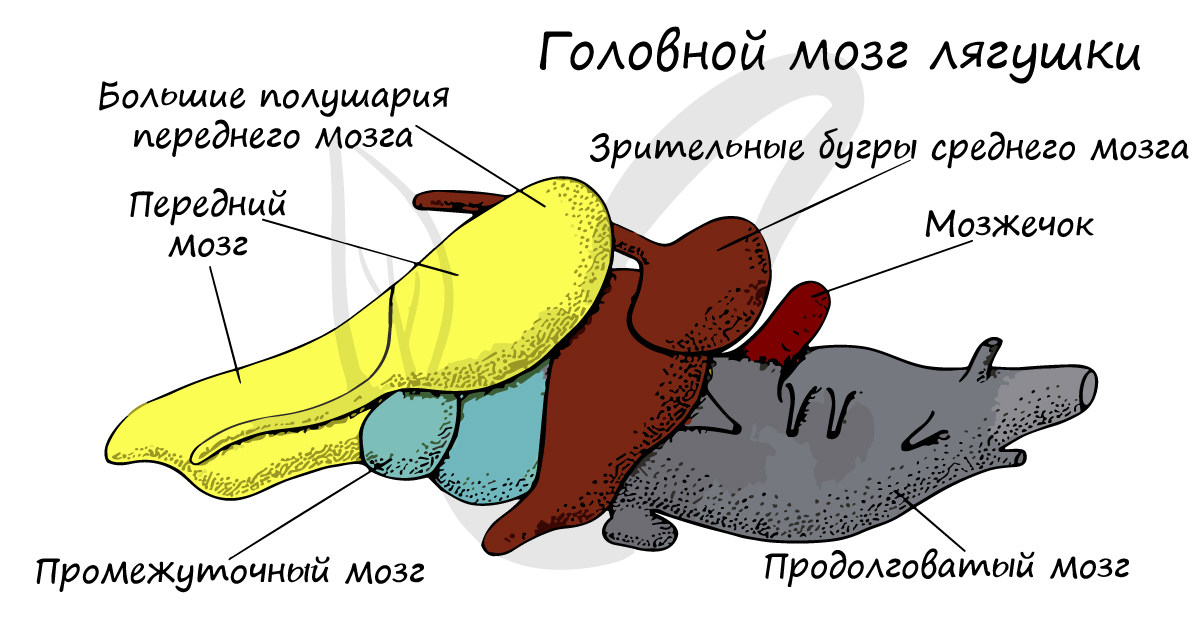
Головной мозг характеризуется рядом прогрессивных черт. Это выражается в относительно более крупных, чем у рыб, размерах переднего мозга, в полном разделении его полушарий и в том, что не только дно боковых желудочков, но и их бока и крыша содержат мозговое вещество. Таким образом, у земноводных имеется настоящий мозговой свод — архипаллиум, который из костных рыб свойствен только двоякодышащим. Средний мозг сравнительно небольших размеров. Мозжечок очень мал, а у некоторых хвостатых (у протеев) он практически незаметен. Слабое развитие этой части головного мозга находится в связи с крайне однообразными, несложными движениями, совершаемыми амфибиями. От головного мозга отходят десять нар головных нервов (I—X), одиннадцатая пара (добавочный нерв) не развита, а двенадцатая отходит за пределами черепа.
Спинномозговые нервы у хвостатых и бесхвостых образуют хорошо выраженные плечевое и поясничное сплетения. Хорошо развита симпатическая нервная система, представленная в основном двумя нервными стволами, расположенными по бокам позвоночника земноводных.
Органы зрения
Глаза амфибий имеют ряд особенностей, связанных с полуназемным образом жизни. Последнее выражается: 1) в наличии подвижных век, защищающих глаза от высыхания и загрязнения; при этом, кроме верхнего и нижнего века, имеется еще третье веко, или мигательная перепонка, расположенная в переднем углу глаза; 2) в выпуклой (а не плоской, как у рыб) форме роговицы и линзовидной (а не круглой, как у рыб) форме хруст лика; обе последние особенности определяют более дальнозоркое зрение амфибий (интересно, что при нахождении в воде роговица у земноводных становится плоской); 3) в более совершенной аккомодации зрения, достигаемой путем перемещения хрусталика под действием ресничного мускула.
Орган слуха
Орган слуха земноводных сравнительно с таковым рыб устроен значительно более сложно и приспособлен к лучшему восприятию звуковых раздражений в воздушной среде. Наиболее полно это выражено у высших (бесхвостых) амфибий. Кроме внутреннего уха, представленного, как и у рыб, перепончатым лабиринтом, у земноводных имеется еще среднее ухо. Последнее представляет полость, один конец которой открывается в ротоглотку, а другой подходит к самой поверхности головы и затянут тонкой перепонкой, известной под названием барабанной. Полость эта образует изгиб, вершина которого находится у края перепончатого лабиринта. Верхняя часть полости от барабанной перепонки до перепоночного лабиринта носит название барабанной полости. В ней расположена палочковидная косточка (stapes), которая одним концом упирается в овальное окно внутреннего уха, другим — в барабанную перепонку. Нижняя часть полости среднего уха, открывающаяся в ротоглотку, именуется евстахиевой трубой.
Данные сравнительной анатомии и эмбриологии показывают, что полость среднего уха гомологична брызгальцу рыб, т. е. рудиментарной жаберной щели, лежащей между челюстной и подъязычной дугами, а слуховая косточка гомологична верхнему отделу подъязычной дуги, т. е. гиомандибуляре. На этом примере видно, что важное изменение органа может достигаться не только путем появления новых образований, но и путем видоизменения и смены функций у образований, имевшихся и ранее.
У безногих и хвостатых барабанная перепонка и барабанная полость отсутствуют, но слуховая косточка хорошо развита. Редукция среднего уха у этих групп, видимо, явление вторичное.
Органы обоняния амфибий
Органы обоняния амфибий представляют парные обонятельные капсулы, которые сообщаются с наружной средой парными же наружными ноздрями, от обонятельных капсул отходят внутренние ноздри ( хоаны), сообщающие их с ротоглоточной полостью. У амфибий, как и у всех наземных позвоночных, указанная система служит не только для восприятия запахов, но и для дыхания.
Органы боковой линии свойственны личинкам всех амфибий. Во взрос-лом состоянии они сохраняются только у водных форм хвостатых амфибий и немногих, тоже водных, бесхвостых. Однако, в отличие от рыб, чувствующие клетки этого органа расположены не в углубленном канале, а лежат поверхностно в коже.
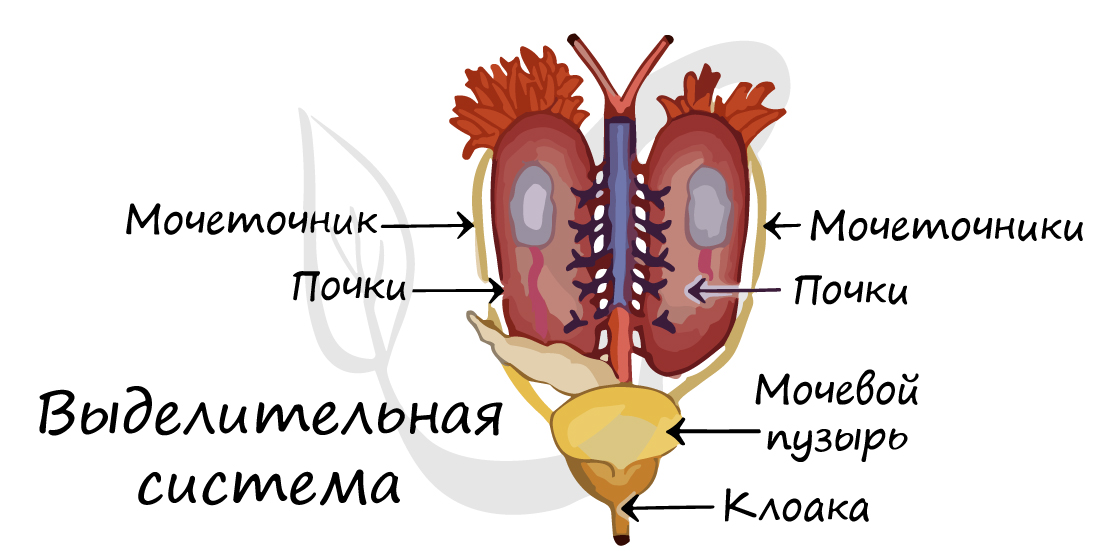
Мочеполовые органы
Мочеполовые органы земноводных устроены по типу мочеполовых органов хрящевых рыб. Органом выделения в зародышевом состоянии служит пронефрос; у взрослых — мезонефрос с его типичным выводным путем — вольфовым каналом. Мочеточники открываются в клоаку. Сюда же у высших наземных земноводных открывается мочевой пузырь. После его наполнения моча через то же отверстие выводится в клоаку и далее изгоняется наружу.
Основным продуктом белкового обмена у амфибий служит мочевина, которая не очень токсична, но для выведения из организма требует большого количества воды, в которой она растворяется. Физиологически это вполне оправдано, так как восприятие воды организмом у амфибий в подавляющем большинстве случаев не встречает затруднений.
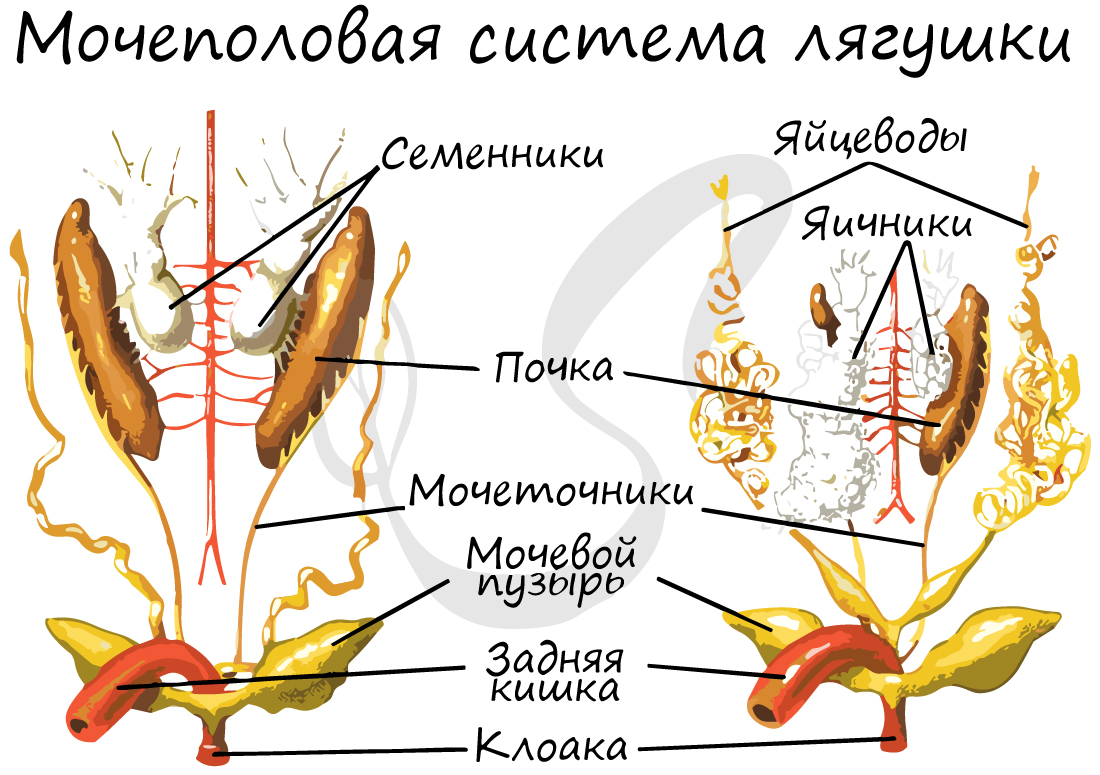
Парные семенники не имеют самостоятельных выводных путей. Семявыносящие канальцы проходят через передний отдел почки и впадают в вольфов канал, который, таким образом, служит не только мочеточником, но и семявыносящим каналом. Каждый вольфов канал у самцов перед впадением в клоаку образует расширение — семенной пузырек, служащий для временного резервирования семени. Над семенниками лежат жировые тела — образования неправильной формы, желтого цвета. Они служат для питания семенников и развивающихся в них сперматозоидов. Величина жировых тел меняется по сезонам. Осенью они велики, весной же, во время интенсивного сперматогенеза, вещество их энергично расходуется и размеры резко сокращаются. Копулятивных органов у подавляющего большинства амфибий нет.
Яичники парные; над ними также лежат жировые тела. Созревшие яйца попадают в полость тела, откуда они поступают в воронкообразные расширения парных яйцеводов — мюллеровы каналы. Яйцеводы — длинные, сильно извитые трубки,



