Строение и функции ядра
Вопрос 1. Каковы функции ядра клетки?
Ядро в клетке выполняет основные функции:
1. хранение и воспроизведение наследственной информации, которая хранится в ядре в виде молекул ДНК, входящих в состав хромосом;
2. регуляция обмена веществ в клетке осуществляется благодаря тому, что в ядре содержится наследственная информация о строении клеточных белков в составе ядерных хромосом.
Вопрос 2. Какие организмы относятся к прокариотам?
Прокариоты — это организмы, клетки которых не имеют оформленного ядра. К ним относят бактерии, сине-зеленые водоросли (цианобактерии) и археи.
Вопрос 5. Каковы функции ядрышек?
Ядрышки – это округлые, сильно уплотненные, не ограниченные мембраной участки ядра. Форма их, размеры и количество зависит от функционального состояния ядра. В клетке, выполняющей функцию синтеза большого количества белка, в ядре будет несколько ядрышек или они будут крупные и рыхлые, т.е. функция ядрышка – это синтез рРНК и сборка малой и большой субъединиц рибосом. В составе ядрышка находится: 80% белка, 10-15% РНК, небольшое количество ДНК и другие химические компоненты. В профазу деления клетки субъединицы рибосом через ядерные поры выходят в цитоплазму, ДНК ядрышка упаковывается на хромосомы, имеющие вторичную перетяжку или ядрышковый организатор, и соответственно, ядрышко как структура распадается и становится не видимой структурой, поэтому иногда говорят, что оно «растворяется».
Вопрос 6. Из чего состоит хромосома?
Хромосома представляет собой молекулу ДНК, соединенную с особым белком, придающим ей компактность.
Вопрос 7. Где располагаются хромосомы у бактерий?
В клетках бактерий нет оформленного ядра. Генетический аппарат бактерий представлен одной кольцевой молекулой ДНК (бактериальной хромосомой), которая присоединена в определенном месте к клеточной мембране и занимает в цитоплазме пространство, называемое нуклеоидом.
Вопрос 9. Как называется набор хромосом в соматических клетках?
Как правило, соматические клетки содержат двойной набор хромосом, который называется диплоидным.
Вопрос 10. Какой набор хромосом в гаметах?
Гаметы содержат только по одной хромосоме каждого вида, т. е. имеют одинарный набор хромосом, который называется гаплоидным.
Вопрос 11. Какой гаплоидный набор хромосом в клетках рака, если диплоидный равен 118?
Если диплоидный набор хромосом в клетках равен 118, то гаплоидный будет в два раза меньше — 59 (118/2=59).
Вопрос 12. Может ли диплоидный набор содержать нечетное число хромосом?
Диплоидный набор хромосом может содержать нечетное количество хромосом. Существуют организмы, у которых в соматических клетках имеется только одна половая хромосома. Например, у некоторых насекомых (клопы, кузнечики) самки гомогаметны (XX), а самцы имеют только одну половую хромосому (ХО).
Хромосомы бактерий

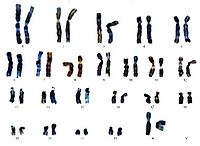
Хромосо́мы (греч. χρώμα — цвет и греч. σώμα — тело) — хорошо окрашиваемые включения в ядре эукариотической клетки, которые становятся легко заметными в определённых фазах клеточного цикла (во время митоза или мейоза). Хромосомы представляют собой высокую степень конденсации хроматина, постоянно присутствующего в клеточном ядре. Исходно термин был предложен для обозначения структур, выявляемых в эукариотических клетках, но в последние десятилетия все чаще говорят о бактериальных хромосомах. В хромосомах сосредоточена большая часть наследственной информации.
Содержание
Хромосомы эукариот
Хромосомы эукариот имеют сложное строение. Основу хромосомы составляет линейная (не замкнутая в кольцо) макромолекула дезоксирибонуклеиновой кислоты (ДНК) значительной длины (например, в молекулах ДНК хромосом человека насчитывается от 50 до 245 миллионов пар азотистых оснований). В растянутом виде длина хромосомы человека может достигать 5 см. Помимо неё, в состав хромосомы входят пять специализированных белков — H1, H2A, H2B, H3 и H4 (так называемые гистоны) и ряд негистоновых белков. Последовательность аминокислот гистонов высококонсервативна и практически не различается в самых разных группах организмов.
В интерфазе хроматин не конденсирован, но и в это время его нити представляют собой комплекс из ДНК и белков. Макромолекула ДНК обвивает октомеры (структуры, состоящую из восьми белковых глобул) гистоновых белков H2A, H2B, H3 и H4, образуя структуры, названные нуклеосомами. В целом вся конструкция несколько напоминает бусы. Последовательность из таких нуклеосом, соединённых белком H1, называется нуклеофиламентом (nucleofilament), или нуклеосомной нитью, диаметром около 10 нм.
В ранней интерфазе (фаза G1) основу каждой из будущих хромосом составляет одна молекула ДНК. В фазе синтеза (S) молекулы ДНК вступают в процесс репликации и удваиваются. В поздней интерфазе (фаза G2) основа каждой из хромосом состоит из двух идентичных молекул ДНК, образовавшихся в результате репликации и соединённых между собой в районе центромерной последовательности.
Перед началом деления клеточного ядра хромосома, представленная на этот момент цепочкой нуклеосом, начинает спирализовываться, или упаковываться, образуя при помощи белка H1 более толстую хроматиновую нить, или хроматиду, (chromatin fiber) диаметром 30 нм. В результате дальнейшей спирализации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, по-прежнему соединены между собой в районе центромеры (подробнее о судьбе хромосом при клеточном делении см. статьи митоз и мейоз).
Центромера
Первичная перетяжка
X. п., в которой локализуется центромера и которая делит хромосому на плечи.
Вторичные перетяжки
Морфологический признак, позволяющий идентифицировать отдельные хромосомы в наборе. От первичной перетяжки отличаются отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. У человека это 13, 14, 15, 21 и 22 хромосомы.
Типы строения хромосом
Различают четыре типа строения хромосом:
Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода.
Спутники (сателлиты)
Сателлит — это округлое или удлинённое тельце, отделённое от основной части хромосомы тонкой хроматиновой нитью, по диаметру равный или несколько меньший хромосоме. Хромосомы, обладающие спутником принято обозначать SAT-хромосомами. Форма, величина спутника и связывающей его нити постоянны для каждой хромосомы.
Зона ядрышка
Зоны ядрышка (организаторы ядрышка) — специальные участки, с которыми связано появление некоторых вторичных перетяжек.
Хромонема
Хромонема — это спиральная структура, которую удаётся увидеть в декомпактизованных хромосомах через электронный микроскоп. Впервые наблюдалась Баранецким в 1880 году в хромосомах клеток пыльников традесканции, термин ввёл Вейдовский. Хромонема может состоять из двух, четырёх и более нитей, в зависимости от исследуемого объекта. Эти нити образуют спирали двух типов:
Хромосомные перестройки
Нарушение структуры хромосом происходит в результате спонтанных или спровоцированных изменений (например, после облучения).
Гигантские хромосомы
Такие хромосомы, для которых характерны огромные размеры, можно наблюдать в некоторых клетках на определённых стадиях клеточного цикла. Например, они обнаруживаются в клетках некоторых тканей личинок двукрылых насекомых (политенные хромосомы) и в ооцитах различных позвоночных и беспозвоночных (хромосомы типа ламповых щёток). Именно на препаратах гигантских хромосом удалось выявить признаки активности генов.
Политенные хромосомы
Впервые обнаружены Бальбиани в 1881-го, однако их цитогенетическая роль была выявлена Костовым, Пайнтером, Гейтцем и Бауером. Содержатся в клетках слюнных желёз, кишечника, трахей, жирового тела и мальпигиевых сосудов личинок двукрылых.
Хромосомы типа ламповых щеток
Обнаружены Рюккертом в 1892 году. По длине превышают политенные хромосомы, наблюдаются в ооцитах на стадии первого деления мейоза, во время которой процессы синтеза, приводящие к образованию желтка, наиболее интенсивны. Общая длина хромосомного набора в ооцитах некоторых хвостатых амфибий достигает 5900 мкм.
Бактериальные хромосомы
Прокариоты (архебактерии и бактерии, в том числе митохондрии и пластиды, постоянно обитающие в клетках большинства эукариот) не имеют хромосом в собственном смысле этого слова. У большинства из них в клетке имеется только одна макромолекула ДНК, замкнутая в кольцо (эта структура получила название нуклеоид). У ряда бактерий обнаружены линейные (не замкнутые в кольцо) макромолекулы ДНК. Помимо нуклеоида или линейных макромолекул, ДНК может присутствовать в цитоплазме прокариотных клеток в виде небольших замкнутых в кольцо молекул ДНК, так называемых плазмид, содержащих обычно незначительное, по сравнению с бактериальной хромосомой, число генов. Состав плазмид может быть непостоянен, бактерии могут обмениваться плазмидами в ходе парасексуального процесса.
Имеются данные о наличии у бактерий белков, связанных с ДНК нуклеоида, но гистонов у них не обнаружено.
Генетика бактерий. Наследственный аппарат, изменчивость
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
Генетика бактерий
1.Важнейшими признаками живых организмов являются изменчивость и наследственность.
Основу наследственного аппарата бактерий, как и всех других организмов, составляет ДНК (у РНК-содержащих вирусов — РНК).
Наряду с этим наследственный аппарат бактерий и возможности его изучения имеют ряд особенностей:
бактерии — гаплоидные организмы, т. е. они имеют 1 хромосому. В связи с этим при наследовании признаков отсутствует явление доминантности;
• бактерии обладают высокой скоростью размножения, в связи с чем за короткий промежуток времени (сутки) сменяется несколько десятков поколений бактерий. Это дает возможность изучать огромные по численности популяции и достаточно легко выявлять даже редкие по частоте мутации. Наследственный аппарат бактерий представлен хромосомой. У бактерий она одна. Если и встречаются клетки с 2, 4 хромосомами, то они одинаковые.
Хромосома бактерий — это молекула ДНК. Длина этой молекулы достигает 1,0 мм и, чтобы “уместиться” в бактериальной клетке, она не линейная, как у эукариотов, а суперспирализо-вана в петли и свернута в кольцо. Это кольцо в одной точке прикреплено к цитоплазматической мембране. На бактериальной хромосоме располагаются отдельные гены. У кишечной палочки, например, их более 2 тыс.
2. Генотип (геном) бактерий представлен не только хромосомными генами. Функциональными единицами генома бактерий, кроме хромосомных генов, являются:
IS-последовательности — короткие фрагменты ДНК. Они не несут структурных (кодирующих тот или иной белок) генов, а содержат только гены, ответственные за транспозицию (способность IS-последовательностей перемещаться по хромосоме и встраиваться в различные ее участки). IS-последовательности одинаковы у разных бактерий. Транспозоны — это молекулы ДНК, более крупные, чем IS-последовательности. Помимо генов, ответственных за транспозицию, они содержат и структурный ген, кодирующий тот или иной признак.
Транспозоны легко перемещаются по хромосоме. Их положение сказывается на экспрессии как их собственных структурных генов, так и соседних хромосомных. Транспозоны могут существовать и вне хромосомы, автономно, но неспособны к автономной репликации.
Плазмиды — кольцевые суперспиралевидные молекулы ДНК. Их молекулярная масса колеблется в широких пределах и может быть в сотни раз больше, чем у транспозонов.
Плазмиды содержат структурные гены, наделяющие бактериальную клетку разными, весьма важными для нее свойствами:
• R-плазмиды — лекарственной устойчивостью;
• Col-плазмиды — способностью синтезировать колицины;
• F-плазмиды — передавать генетическую информацию;
• Шу-плазмиды — синтезировать гемолизин;
• Тох-плазмиды — синтезировать токсин;
• плазмиды биодеградации — разрушать тот или иной субстрат и т. д.
Многие плазмиды имеют в своем составе гены трансмиссивности и способны передаваться от одной клетки к другой при конъюгации (обмене генетической информацией). Такие плазмиды называются трансмиссивными.
3. Наличие F-плазмиды (фактор фертилъности, половой фактор)
придает бактериям функции донора, и такие клетки способны передавать свою генетическую информацию другим, F-клеткам. Можно сказать, что наличие F-плазмиды является фенотипиче-ским выражением (проявлением) пола у бактерий: с F-плазмидой связана не только донорская функция, но и некоторые другие фенотипические признаки — наличие F-пилей (половых ресничек) и чувствительность к L-фагам. С помощью F-ресничек устанавливается контакт между донорскими и реципиентными клетками. Через их канал и передается донорская ДНК при рекомбинации. На половых ресничках расположены рецепторы для мужских fj-фагов. F-клетки не имеют таких рецепторов и нечувствительны к таким фагам.
4. У бактерий различают 2 вида изменчивости — фенотипическую и генотипическую.
Фенотипическая изменчивость — модификация — не затрагивает генотип, но затрагивает большинство особей популяции. Модификации не передаются по наследству и с течением времени затухают, т. е. возвращаются к исходному фенотипу через большее (длительные модификации) или меньшее (кратковременные модификации) число поколений.
Генотипическая изменчивость затрагивает генотип. В ее основе лежат мутации и рекомбинации.
Мутации бактерий принципиально не отличаются от мутаций эукариотических клеток. Особенностью мутаций у бактерий является относительная легкость их выявления, так как имеется возможность работать с большими по численности популяциями бактерий. По происхождению мутаиии могут быть:
• индуцированными. По протяженности:
• хромосомными. По направленности:
Рекомбинации (обмен генетическим материалом) у бактерий отличаются от рекомбинаций у эукариот:
• у бактерий имеется несколько механизмов рекомбинаций;
• при рекомбинациях у бактерий образуется не зигота, как у эукариот, а мерозигота (несет полностью генетическую информацию реципиента и часть генетической информации донора в виде дополнения);
• у бактериальной клетки-рекомбината изменяется не только качество, но и количество генетической информации. Трансформация — это обмен генетической информацией у бактерий путем введения в бактериальную клетку-реципиент готового препарата ДНК (специально приготовленного или непосредственно выделенного из клетки-до нора). Чаще всего передача генетической информации происходит при культивировании реципиента на питательной среде, содержащей ДНК донора. Для восприятия донорской ДНК при трансформации клетка-реципиент должна находиться в определенном физиологическом состоянии (компетентности), которое достигается специальными методами обработки бактериальной популяции.
При трансформации передаются единичные (чаще 1) признаки. Трансформация является самым объективным свидетельством связи ДНК или ее фрагментов с тем или иным фенотипическим признаком, поскольку в реципиентную клетку вводится чистый препарат ДНК.
Трансдукция — обмен генетической информацией у бактерий путем передачи ее от донора к реципиенту с помощью умеренных (трансдуцирующих) бактериофагов.
Трансдуцирующие фаги могут переносить 1 или более генов (признаков). Трансдукиия бывает:
• специфической — переносится всегда один и тот же ген;
• неспецифической — передаются разные гены.
Это связано с локализацией трансдуиируюших фагов в геноме донора:
• в случае специфической трансдукции они располагаются всегда в одном месте хромосомы;
• при неспецифической их локализация непостоянна. Конъюгация — обмен генетической информацией у бактерий путем передачи ее от донора к реципиенту при их прямом контакте. После образования между донором и реципиентом конъюга-ционного мостика одна нить ДНК-донора поступает по нему в клетку-реципиент. Чем дольше контакт, тем большая часть донорской ДНК может быть передана реципиенту.
Основываясь на прерывании конъюгации через определенные промежутки времени, можно определить порядок расположения генов на хромосоме бактерий — построить хромосомные карты бактерий (произвести картирование бактерий).
Какой набор хромосом у бактерий
• В клетках большинства прокариот присутствует одна кольцевая хромосома
• Генетическая гибкость прокариот и их адаптационные возможности усиливаются при заражении бактериофагами и при переносе плазмид
• Быстрая эволюция генома прокариот обеспечивается транспозонами и другими мобильными элементами
У большинства прокариот присутствует одна кольцевая хромосома, и они являются гаплоидными клетками. В следующем разделе рассмотрены вопросы организации хромосомы в дискретную структуру, нуклеоид. В настоящее время мы располагаем данными секвенирования более 200 геномов прокариот, размеры которых варьируют от 580 кнп (Mycoplasma genitalium) до 9 Мнп (Streptomyces, Myxococcus).
Геном таких хорошо известных бактерий, как Е. coli и В. subtilis, занимает среднее положение (4-5 Мнп). Относительно небольшие размеры прокариотических хромосом, по сравнению с хромосомами высших эукариот, объясняются их компактностью и небольшим количеством некодирующих последовательностей ДНК. В общем, гены, необходимые для постоянного роста и поддержания жизнеспособности прокариот, находятся в хромосоме, а генетическая гибкость микроорганизмов обеспечивается различными мобильными элементами.
У некоторых бактерий хромосома имеет линейную форму, или же в клетке содержится несколько таких хромосом. Встречаются виды, для которых характерны обе эти особенности. Например, клетки Streptomyces содержат линейную хромосому. Концы этой хромосомы замкнуты белковым мостиком, и это объясняет, почему долгое время на генетической карте этих бактерий изображали круговую хромосому.
Бактерии Rhodobacter sphaeroides обладают двумя большими кольцевыми хромосомами (3,0 и 0,9 Мнп), в каждой из которых присутствует много необходимых генов домашнего хояйства. Возбудитель Лайм-боррелиоза, бактерия Воrrelia burgdorferi характеризуется наличием нескольких линейных хромосом.
Стабильные элементы внехромосомной ДНК, которые не несут необходимых генов домашнего хозяйства и поэтому не являются существенными, называются плазмиды. На рисунке ниже перечислены некоторые хорошо изученные плазмиды бактерий. Эти данные дают представление о размерах генов, которые могут нести эти плазмиды. Плазмиды обычно невелики, варьируя по размерам от 2 до 1000 кнп, и обладают кольцевой структурой.
Так же как и в случае хромосом, известны исключения, и некоторые крупные плазмиды достигают размера 1 Мнп или больше, и небольшая их часть обладает линейной структурой. Все плазмиды несут гены, управляющие их репликацией, обычно включающей различные элементы аппарата репликации клетки хозяина. К числу важнейших генов, которые несут плазмиды, относятся гены устойчивости к антибиотикам и обусловливающие патогенные свойства или деградацию необычных источников углерода.
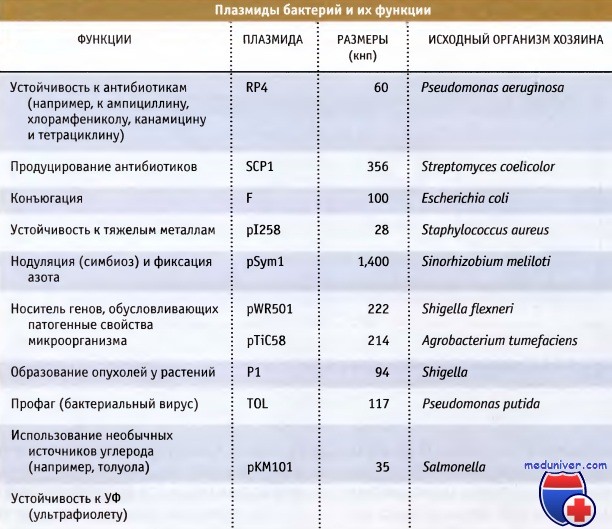 Плазмиды бактерий и их функции.
Плазмиды бактерий и их функции.
Плазмиды могут распространяться между организмами различными путями. Конъюгация представляет собой процесс, напоминающий спаривание, при котором присходит прямой перенос ДНК от клетки донора, содержащей плазмиду, в клетку реципиента. В донорской плазмиде закодированы функции, необходимые дляУ большинства прокариот присутствует одна кольцевая хромосома, и они являются гаплоидными клетками. В следующем разделе рассмотрены вопросы организации хромосомы в дискретную структуру, нуклеоид.
В настоящее время мы располагаем данными секвенирования более 200 геномов прокариот, размеры которых варьируют от 580 кнп (Mycoplasma genitalium) до 9 Мнп (Streptomyces, Myxococcus). Геном таких хорошо известных бактерий, как Е. coli и В. subtilis, занимает среднее положение (4-5 Мнп). Относительно небольшие размеры прокариотических хромосом, по сравнению с хромосомами высших эукариот, объясняются их компактностью и небольшим количеством некодирующих последовательностей ДНК. В общем, гены, необходимые для постоянного роста и поддержания жизнеспособности прокариот, находятся в хромосоме, а генетическая гибкость микроорганизмов обеспечивается различными мобильными элементами.
У некоторых бактерий хромосома имеет линейную форму, или же в клетке содержится несколько таких хромосом. Встречаются виды, для которых характерны обе эти особенности. Например, клетки Streptomyces содержат линейную хромосому. Концы этой хромосомы замкнуты белковым мостиком, и это объясняет, почему долгое время на генетической карте этих бактерий изображали круговую хромосому.
Бактерии Rhodobacter sphaeroides обладают двумя большими кольцевыми хромосомами (3,0 и 0,9 Мнп), в каждой из которых присутствует много необходимых генов домашнего хояйства. Возбудитель Лайм-боррелиоза, бактерия Воrrelia burgdorferi характеризуется наличием нескольких линейных хромосом.
Стабильные элементы внехромосомной ДНК, которые не несут необходимых генов домашнего хозяйства и поэтому не являются существенными, называются плазмиды. На рисунке ниже перечислены некоторые хорошо изученные плазмиды бактерий. Эти данные дают представление о размерах генов, которые могут нести эти плазмиды. Плазмиды обычно невелики, варьируя по размерам от 2 до 1000 кнп, и обладают кольцевой структурой.
Так же как и в случае хромосом, известны исключения, и некоторые крупные плазмиды достигают размера 1 Мнп или больше, и небольшая их часть обладает линейной структурой. Все плазмиды несут гены, управляющие их репликацией, обычно включающей различные элементы аппарата репликации клетки хозяина. К числу важнейших генов, которые несут плазмиды, относятся гены устойчивости к антибиотикам и обусловливающие патогенные свойства или деградацию необычных источников углерода.
Плазмиды могут распространяться между организмами различными путями. Конъюгация представляет собой процесс, напоминающий спаривание, при котором присходит прямой перенос ДНК от клетки донора, содержащей плазмиду, в клетку реципиента. В донорской плазмиде закодированы функции, необходимые для обеспечения контакта с клеткой реципиентом, инициации переноса реплицированной ДНК, и самого переноса ДНК в клетку реципиента.
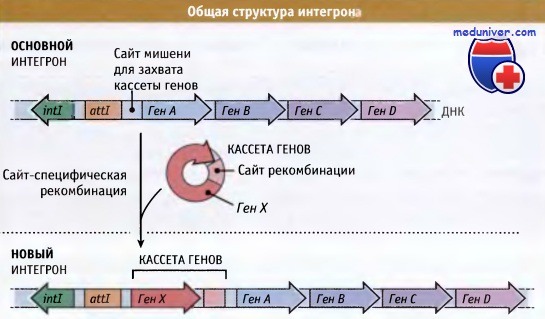 Структура типичного интегрона после накопления нескольких кассет генов.
Структура типичного интегрона после накопления нескольких кассет генов.
Плазмиды также могут передаваться при прямом захвате ДНК (трансформация) или с участием бактериофага (трансдукция). Независимо от наличия экстрахромосомных элементов, изменения в бактериальном геноме могут возникать за счет процессов гомологичной и сайт-специфической рекомбинации.
Для многих бактерий важными источниками генетической вариабельности, по-видимому, являются бактериофаги (бактериальные вирусы). Результаты секвенирования показывают, что в геноме многих бактерий присутствуют интегрированные последовательности бактериофага (профаги). Например, у Е. coli их по меньшей мере 9, а у B. subtilis 10. Некоторые из этих профагов обладают дефектами, например у них присутствуют делеции и другие мутации, что делает маловероятным их дальнейшую активацию с образованием инфекционных бактериофагов.
В некоторых случаях профаги несут гены, которые обеспечивают некоторые преимущества клеткам хозяина: это относится к системам рестрикции и модификации, устойчивости к УФ и к таким детерминантом патогенности, как токсины.
Наконец, бактериальный геном также содержит много мобильных генетических элементов, которые распространяются посредством транспозиции. Инсерционные последовательности содержат минимальные элементы, которые, в своей простейшей форме, вместе с фланкирующими последовательностями, обладают только одним геном транспозазы. При инициации транспозиции, фланкирующие последовательности узнаются белком транспозазой.
Затем происходит мобилизация ферментов клетки хозяина, участвующих в репликации и репарации ДНК. Они завершают процесс вставки последовательности в необходимом месте. Более сложные транспозоны несут дополнительные гены, способные обеспечить адаптационные преимущества клеткам хозяина. Наиболее известными примерами транспозонов у бактерий являются гены устойчивости к антибиотикам, однако подобным образом могут переноситься также многие другие гены. Близкие к транспозонам элементы могут катализировать несколько таких типов перегруппировок ДНК, как инверсии и делеции.
Интегроны представляют собой особенно важный инструмент адаптивной перегруппировки генома. Как показано на рисунке ниже, интегроны обычно состоят из гена интегразы, примыкающего к нему сайта мишени для захвата кассеты генов, и сильного промотора, регулирующего экспрессию захваченных генов. В кассетах, часто включающих гены устойчивости к антибиотикам, содержатся последовательности, которые позволяют им включаться на место мишени под действием белка интегразы. Интегроны могут расти за счет последовательного захвата различных кассет генов, что способствует быстрому развитию таких бактерий, которые устойчивы к ряду антибиотиков.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в содержание раздела «генетика» на нашем сайте



