Какой компонент клеточной стенки является обязательным для грамположительных и грамотрицательных
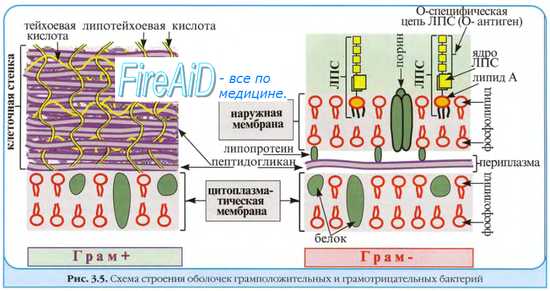
Грамположительные бактерии имеют сравнительно просто организованную, но мощную клеточную стенку. Она состоит преимущественно из множества слоев пептидогликана, составляющего до 90% её сухой массы (см. рис. 4-4), часто включающих вместо диаминопимелиновой кислоты лизин и уникальные водорастворимые полимеры тейхоевых кислот, состоящих из 8-50 остатков глицерина или рибита, связанных между собой фосфодиэфирными связями. Тейхоевые кислоты [от греч. teichos, стенка] могут составлять до 50% сухого веса клеточной стенки. Известно два типа кислот — рибиттейхоевые (состоят из остатков рибитфосфата и 10-50 остатков спирта) и глицеринтейхоевые (состоят из остатков глицерофосфата и 20 остатков спирта). Клеточная стенка каждого вида содержит только один тип теихоевых кислот (за исключением вида Streptomyces). Тейхоевые кислоты — основные поверхностные Аг многих бактерий. У большей части грамноложительных бактерий также имеются периплазматические теихоевые кислоты, располагающиеся между клеточной стенкой и ЦПМ. Периплазматические кислоты, выявляемые горячей кислотной экстракцией, являются группоспецифичными Аг.
Клеточная стенка грамположительных бактерий не содержит ЛПС, но может включать различные белки. Содержание последних весьма вариабельно. Для некоторых бактерий (например, стрептококков группы А) белки служат серовароспецифичными Аг.
Аутолизины бактерий. Клеточные стенки бактерий содержат аутолизины — ферменты, растворяющие пептидогликановый слой. Их активность необходима для процессов роста клеточной стенки, деления клеток, споруляции и достижения состояния компетентности при трансформации (см. ниже).
Удаление клеточной стенки, защищающей прилежащую ЦПМ, приводит к лизису бактерии либо к образованию протопластов или сферопластов (из грамположительных или грамотрицательных бактерий соответственно). Бактерии, лишённые клеточной стенки, лишь в изотонической среде способны поглощать 02 и выделять С02, а также размножаться.
Сферопласты более устойчивы к изменениям осмотического давления и длительно сохраняются в неизотонической среде.
Какой компонент клеточной стенки является обязательным для грамположительных и грамотрицательных
Грамотрицателъные бактерии имеют сравнительно тонкую клеточную стенку. В ней выделяют два слоя — пластичный и ригидный. Последний образован одним, редко двумя слоями пептидогликана, содержание которого составляет не более 20% сухой массы клеточной стенки. На пептидогликановом каркасе расположены фосфолипиды, липополисахариды (ЛПС) и белки, образующие пластичный слой. Толщина пластичного слоя значительно превышает размеры монослоя пептидогликана. Его компоненты расположены мозаично и могут образовывать дополнительную внешнюю мембрану либо переходить в капсулу.
• Фосфолипиды клеточной стенки пластичного слоя прикреплены к пептидогликану липопротеинами, пересекающими периплазматическое пространство. Обработка детергентами (например, додецилсульфатом натрия) приводит к нарушению этих связей. Основное отличие внешнего фосфолипидного слоя от внутреннего ригидного — высокое содержание липополисахариды.
• Липополисахариды клеточной стенки состоят из липидной части (липид А), базисной части молекулы полисахарида (сердцевина) и боковых полисахаридных цепей (рис. 4-5). Иммуногенные свойства проявляют боковые полисахаридные цепи и сердцевина. Боковые полисахаридные цепи отвечают за антигенную специфичность молекулы липополисахаридов и называются О-Аг. Липидная часть термоустойчива и отвечает за биологические эффекты ЛПС. Структура ЛПС имеет большое диагностическое значение, поскольку разные виды или серовары патогенных грамотрицательных бактерий отличаются друг от друга составом боковых цепей ЛПС внешней мембраны.
• Белки, входящие в состав пластичного слоя, подразделяют (в зависимости от выполняемых функций) на основные (мажорные) и второстепенные (минорные). К мажорным белкам относят порины, образующие трансмембранные каналы, вовлечённые в транспорт ионов и гидрофильных соединений из внешней среды в периплазму. Минорные белки также могут участвовать в транспорте веществ через пластичный слой (путём облегчённой диффузии или активного транспорта молекул). Некоторые белки играют роль рецепторов для вирусов бактерий и бактериоцинов, а также для донорских пилей при конъюгации.
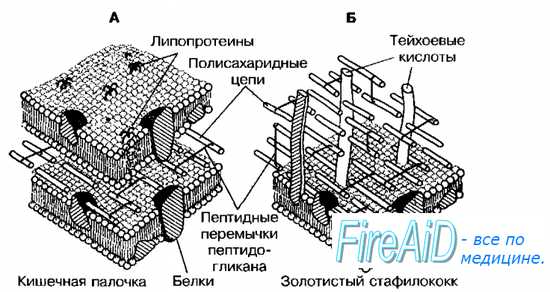 Рис. 4-4. Клеточная стенка грамотрицательных (А) и грамположительных (Б) бактерий.
Рис. 4-4. Клеточная стенка грамотрицательных (А) и грамположительных (Б) бактерий.
Внешняя мембрана не пропускает молекулы с большой молекулярной массой, что можно рассматривать как фактор неспецифической устойчивости бактерий к некоторым антимикробным препаратам.
ГРАМОТРИЦА́ТЕЛЬНЫЕ И ГРАМПОЛОЖИ́ТЕЛЬНЫЕ БАКТЕ́РИИ
Том 7. Москва, 2007, стр. 619
Скопировать библиографическую ссылку:
ГРАМОТРИЦА́ ТЕЛЬНЫЕ И ГРАМПОЛОЖ И́ТЕЛЬНЫЕ БАКТЕ́РИИ, отделы бактерий, клетки которых соответственно не окрашиваются или окрашиваются по методу, разработанному дат. врачом-микробиологом Х. К. Грамом (1884). Согласно этому методу, на фиксированные клетки бактерий последовательно воздействуют осно́вными красителями (напр., генциановым фиолетовым) и содержащим иод раствором Люголя. После обработки полученного мазка этанолом грамотрицательные бактерии обесцвечиваются, а грамположительные – сохраняют фиолетовую окраску. Это явление связано с тем, что у грамположительных бактерий после обработки раствором Люголя образуется комплекс иода с генциановым фиолетовым, который не может экстрагироваться из клетки, в то время как у грамотрицательных он легко удаляется при промывании бактерий этанолом, а затем водой. Способность или неспособность бактерий к окрашиванию по Граму обусловлена строением их клеточной стенки. Осн. её компонентом является сложный биополимер пептидогликан (муреин). У грамположительных бактерий клеточная стенка более толстая (20–60 нм) и однородная, с бóльшим, чем у грамотрицательных (до 40 раз), содержанием муреина, который препятствует выходу комплекса иода с красителем из клетки; кроме того, в ней присутствуют специфич. тейховые, липотейховые и тейхуроновые кислоты. У грамотрицательных бактерий стенка (толщина 14–18 нм) состоит из двух и более легко различимых слоёв и покрыта внешней мембраной, образованной белками, фосфолипидами и липополисахаридами и выступающей в качестве барьера, защищающего клетку от проникновения мн. соединений (один из механизмов неспецифич. устойчивости грамотрицательных бактерий к антибиотикам). Способность (или неспособность) окрашиваться по Граму служит важным таксономич. признаком бактерий.
Клеточная стенка бактерии
Bacterial cell wall
Клеточная стенка – обязательный структурный элемент бактериальной клетки, за исключением микоплазм и L-форм. У большинства бактерий клеточная стенка невидима в обычный микроскоп без специальной обработки. У крупных форм, в частности, у серобактерий, стенки заметны отчетливо. При помещении клеток в 1–2 %-ный гипертонический раствор NaCl или раствор глюкозы, происходит пазмолиз и контуры клеточной стенки приобретают четкость. В этом случае она хорошо видна при фазово-контрастной микроскопии.
Строение клеточной стенки
По строению клеточных стенок бактерии делят на две большие группы: грамположительные бактерии и грамотрицательные бактерии. Окраска бактерий по Граму позволяет экспериментально разделить бактерии на эти две группы.
Строение и отличие клеточных стенок грамположительных бактерий и грамотрицательных бактерий рассмотрено в одноименных статьях.
Химический состав клеточной стенки
Основной компонент клеточной стенки большинства бактерий – муреин. Он относится к классу пептидогликанов.
Муреин – это гетерополимер. Его основа –неразветвленные гетерополимерныецепочки, состоящие из чередования остатков N-ацетилглюкозамина и N—ацетилмурамовой кислоты, которые соединены между собой β-1,4-гликозидными связями.
Остатки N-ацетилмурамовой кислоты лактильными группами соединены при помощи пептидной связи с аминокислотами. В составе муреина обнаружены следующие аминокислоты: L-аланин, мезо-диаминопимелиновая кислота и D-аланин, D-глутаминовая кислота. Диаминопимелиновая кислота находится в мезоформе.
У некоторых бактерий мезо-диаминопимелиновая кислота может быть заменена на L- или D-орнитин, или 2,4-диаминомасляную кислоту, или гомосерин, или гидроксилизин. Все аминокислоты, включая мезо-диаминопимелиновую, играют важную роль в формировании межмолекулярных сшивок, поскольку в образовании пептидных связей принимают участие обе аминогруппы. Таким образом, две гетерополимерные цепи муреина связываются между собой и образуют мешкообразную гигантскую молекулу –муреиновы мешок. Эта молекула выполняет функцию опорного каркаса клеточной стенки.
Строение клеточной стенки и ее химический состав являются постоянной характеристикой для определенного вида бактерий. Эти характеристики служат важным диагностическим признаком, использующимся для идентификации бактерий.
Особенности клеточных стенок бактерий и их значение
Клеточные стенки бактерий, в отличие от клеточных стенок эукариот содержат особые структурные элементы:
Данные структурные элементы являются слабым местом бактерий и используются в борьбе с вызываемыми ими инфекциями. Для этого применяют лекарственные препараты, воздействующие только на клеточные стенки бактерий или на процесс их синтеза, но не оказывающие влияния на клетки растений, человека и животных.
Функции клеточной стенки
Клеточная стенка бактерии выполняет ряд важных функций:
Одновременно отмечается, что клеточная стенка у бактерий не является жизненно важной структурой, поскольку может быть удалена в определенных условиях. В этом случае бактериальные клетки существуют в виде сферопластов и протопластов.
Тот случай, когда встречают и провожают по одежке

Разнообразие форм клеток прокариот не является (по крайней мере не всегда) случайным феноменом эволюции этих организмов. Исследования показали, что форма бактерий может быть обусловлена физическими законами среды обитания: в вязкой среде эффективнее перемещаются микрообитатели спиральные формы, а следовать направлению лучше могут изогнутые вибрионы и т.д. Согласно расчетам наиболее удобна для микроскопических одноклеточных прокариот форма палочек, которые благодаря своей форме могут противостоять броуновскому движению в жидкостях, имеют эффективное соотношение поверхности к объему клетки и могут закрепляться на субстрате…Авторы статьи проанализировали исследования эволюции и связи с экологией формы клеток бактерий.
Форма и размер бактериальных клеток, как и свойства их клеточной стенки (что отразилось на широко известном делении бактерий на грамположительных и грамотрицательных) – одни из самых первых признаков, использованных для классификации этих организмов. Разнообразие форм клеток и в то же время постоянство формы клеток на видовом уровне (за некоторым обсуждаемом ниже исключением) позволили довольно подробно и точно определять таксономическую принадлежность бактерий. Однако причины возникновения разнообразия формы и ее стабильность внутри разного уровня таксонов прокариот долго оставались загадкой. Новые методы исследований – электронная микроскопия, методы молекулярной биологии и биохимии, а также исследования физических закономерностей и математическое моделирование помогли установить ряд факторов, определяющих внешнее строение бактерий. В обсуждаемой статье авторы представили анализ исследований связи формы клеток бактерий с их экологией и эволюцией.

Несмотря на то, что основными являются три типа клеток бактерий (заглавная иллюстрация) – сферическая, палочковидная и спиральная – специалисты выделяют довольно большое разнообразие других форм (рис. 1). Известно, что бактерии по строению клеточной стенки можно разделить на два типа (рис. 1, 2). Строение оболочки (клеточной стенки бактерий) в значительной степени связано с ее формой. Среди определяющих форму бактерий факторов на данным момент выделяют несколько основных:
— наличие/отсутствие внешней мембраны (у грамотрицательных бактерий);
— относительная толщина пептидогликанового слоя;
— особенности строения продольных пептидных сшивок между гликановыми нитями, ориентированными перпендикулярно длинной оси клетки: у грамотрицательных образуются напрямую, а у грамположительных через дополнительный мостик.

Ряд авторов отмечают, что морфологическое разнообразие грамотрицательных бактерий выше, чем таковое грамположительных (см. рис. 1). Среди грамположительных бактерий преобладают палочки, часто встречаются кокки и нитевидные формы, а вот изогнутые и спиральные формы очень редки. Палочки также преобладают и среди грамотрицательных бактерий, но второе и третье места по распространенности делят изогнутые и спиральные формы. А вот кокки и одноклеточные нитчатые формы среди грамотрицательных бактерий редки, хотя некоторые палочки и спиральные бактерии в определенных условиях могут приобретать округлую форму, например, в стационарной фазе культивирования и при неблагоприятных условиях.
На настоящий момент превалирует представление, что белки цитоскелета, такие как MreB (Murein cluster B) и FtsZ (Filamenting temperature-sensitive mutant Z) гомологи актина и тубулина эукариот, не являются собственно архитектурными элементами формы клеток, а представляют собой нечто похожее на разметку для активации процессов синтеза/разборки клеточной стенки, являясь сайтами прикрепления соответствующих ферментов и регуляторных белков. Экспериментально было показано, что присутствие белка MreB отвечает за палочкообразную форму клетки, а белок FtsZ отвечает за формирование перегородки и других структур во время деления клетки (так называемое Z-кольцо). Представляется, что белок MreB это основной фактор формирования палочковидной формы: он организует в определенных местах клеточной стенки (там, где будут «стенки палочки») синтез пептидогликана (клеточной стенки) после разделения клетки на дочерние и, таким образом, обеспечивает удлинение клеток. У кокков (сферическая форма) этого белка нет, а наращивание клеточной стенки происходит в кольцевой зоне при делении клетки за счет белка FtsZ и других белков, участвующих в делении клетки. Для объяснения формы клеток прокариот еще одним важным белком считается кресцетин CreS. Его наличие в определенной области затормаживает образование клеточной стенки, что приводит к искривлению клетки в результате неравномерного роста. Так могут получаться изогнутые формы. Есть и другие белки-кандидаты (например, бактофилины), претендующие на роль в процессе формообразования у прокариот, однако их функции пока изучены недостаточно.

Кокки. Можно выделить два типа прокариот, имеющих сферическую форму. Одни кокки («собственно» кокки) в течение всего жизненного цикла остаются сферическими. Другие («производные» кокки) – палочки, вибрионы, и др.- приобретают сферическую форму только в неблагоприятных условиях. Как уже говорилось, у подавляющего большинства «собственно» кокков не обнаружен белок MreB (ответственный за палочковидную форму) и сферическая форма приобретается в ходе процессов роста дочерних клеток в зоне деления материнской клетки. «Производные» кокки получают свою сферическую форму другим путем: за счет, так называемого, «редуктивного» деления, когда многократные деления клеток не перемежаются синтезом клеточной стенки в районе стенок (т.е. удлинением). Очевидно, напрашивается вывод, что кокки произошли от палочковидных бактерий в результате потери основного белка MreB, обеспечивающего удлинение стенок.
Чем же выгодно быть сферическим? У сферической формы наименьшее соотношение площади поверхности к объему, это объясняет их малые размеры, потому кокки являются доминирующей группой в микропорах различных типов почв. Это же свойство выгодно при переживании неблагоприятных условий в случае с «производными» кокками. Поскольку шарообразная форма наименее удобна для управляемого движения, кокки, как правило, лишены «органов движения», например, жгутиков. Показано, что сферическая форма позволяет бактериям быстрее распространяться пассивно с током воды, чем бактериям других форм. Эта закономерность объясняет «любовь» кокков образовывать скопления (диплококки – две клетки, стрептококки – нити клеток, стафилококки – гроздья клеток), которые затормаживают пассивное передвижение, а при необходимости агрегация распадается под действием специальных ферментов, разделяющих склеенные между собой клетки (рис. 3).
Палочки. По-видимому, самая удобная (универсальная) для бактерий форма клеток. Большинство исследователей считает палочки исходной в эволюционном плане формой. Подсчитано, что клетки с соотношением длины к диаметру (l/d) около 3.7 испытывают наименьшее сопротивление среды при активном передвижении в жидких средах, более того выгоднее быть длиннее, чем короче, данного соотношения: чтобы испытывать такое же сопротивление среды, как кокки, палочки должны стать в 130 раз длиннее своего диаметра. При соотношении l/d от 3 до 6 наблюдается наибольшая эффективность поглощения питательных веществ из окружающей среды и их внутриклеточного транспорта. Именно таким формам удобно закрепляться на субстрате. Замечено, что очень успешно палочки «собираются» в (печально известные) биопленки.
Многочисленные нитевидные формы это производные палочек, длина стенок которых во много раз превышает диаметр клетки. Нитевидная форма одна из стратегий избегания хищничества со стороны простейших. Длинные, разветвленные формы получают возможность функционально дифференцировать клетку, что способствует более эффективному питанию в случае дефицита определенных элементов питания.
Извитые (спиральные) формы. Бактерии могут становится извитыми разными способами в разных эволюционных линиях прокариот. Например, Helicobacter pylori, вызывающий язву желудка, особыми ферментами (группы Csd) контролируемо разрезает сшивки между нитями в пептидогликановом слое, благодаря чему правильно организованный цилиндр клеточной стенки скручивается в спираль (рис. 4). Интересно, что грамположительные бактерии не имеют ферментов этой группы, к тому же их сшивки между нитями содержат дополнительные (пентаглициновые) мостики, а не сшиты напрямую, как у грамотрицательных бактерий. Эти обстоятельства в некоторой степени объясняют редкость спиральных форм среди грамположительных бактерий.

По-видимому, другой способ скручиваться изобрели Spirochaetae. Сначала было подозрение, что имеющиеся у спирохет жгутики, расположенные в внутреннем пространстве между мембранами (см. рис. 2, межмембранное пространство), ответственны за скручивание клеток. Действительно, было показано, что извитые формы спирохет в виде плоской волны «используют гены» внутренних жгутиков для образования стяжек в нужных местах для придания волнообразной формы клетке. Однако полученные правильно скрученные спиральные мутантные формы без внутренних жгутиков показали, что спиральные спирохеты используют еще какой-то механизм для скручивания. Представляется, что спиральные формы более эффективны при движении в вязкой среде, чем другие формы бактерий.
Изогнутые формы– вибрионы – можно рассматривать как короткие спиральные формы. Но у вибрионов есть покрайней мере еще один способ изогнуться: при помощи «тормозящего» белка кресцетина CreS (см. выше). Ряд исследований показал, что изогнутая форма вибриона способствует активному движению в жидкости и активному поиску лучшего места (хемотаксису).

Помимо общей формы клетки бактерии также могут иметь дополнительные внешние морфологические элементы – жгутики, мембраны, выросты, ножки – отражающие способности прокариот специфически приспосабливаться к определенным условиям жизни, моделируя для себя субнишевые (в экологическом смысле) пространства (рис. 5). Понятно, что целый ряд факторов, таких как свойства среды, способ питания, хищничество со стороны простейших, взаимодействие с субстратом и др. определяют эволюцию формы клеток бактерий. Интересно, что один и тот же тип клеток, как и дополнительных внешних морфологических приспособлений, может обеспечиваться разными структурными элементами оболочки и молекулярными механизмами в ходе эволюции разных таксонов.



