Тетаническое сокращение мышцы




![]()
![]()
При воздействии на мышцу ритмических раздражений высокой частоты наступает сильное и длительное сокращение мышцы, которое называется тетаническим сокращением или тетанусом. Этот термин впервые применил Э. Вебер в 1821 году.
Тетанус может быть зубчатым (при частоте раздражений 20-40 Гц) или сплошным, гладким (при частоте 50 Гц и выше). Амплитуда тетанического сокращения в 2–4 раза выше амплитуды одиночного сокращения при той же силе раздражения.
I 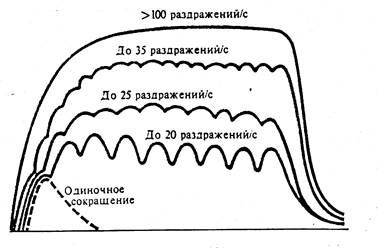
II 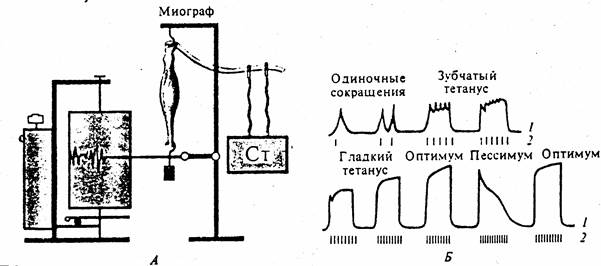
Примечание. Явления пессимума и парабиоза возможны
в условиях эксперимента.
Рис. 81. Формирование тетануса в зависимости от частоты раздражения.
Оптимум и пессимум (по Н.Введенскому). А- схема регистрации; Б- кривые мышечных сокращений (1) при различной частоте раздражений (2).
Гладкий тетанус возникает тогда, когда очередной импульс раздражения действует на мышцу до начала фазы расслабления. При очень большой частоте раздражений каждое очередное раздражение будет попадать на фазу абсолютной рефрактерности и мышца вообще не будет сокращаться. Высота мышечного сокращения при тетанусе зависит от ритма раздражения, а также от возбудимости и лабильности, которые изменяются в процессе сокращения мышцы. Тетанус наиболее высокий при оптимальном ритме, когда каждый последующий импульс действует на мышцу в фазу экзальтации, вызванной предыдущим импульсом. В этом случае создаются наилучшие условия (оптимум силы и частоты раздражения, оптимум ритма) для работы мышцы.
При тетанических сокращениях мышечные волокна утомляются больше, чем при одиночных сокращениях. Поэтому даже в пределах одной мышцы происходит периодическая смена частоты импульсации (вплоть до полного исчезновения) в разных двигательных единицах.
Импульсы с мотонейронов в условиях покоя участвуют в поддержании мышечного тонуса.
Под тонусом понимают состояние естественного постоянного напряжения мышц при невысоких энергетических затратах. В поддержании тонуса участвуют проприорецепторы мышц (мышечные веретена) и центральная нервная система.
Осуществление тонуса скелетных мышц обусловлено функцией медленных двигательных единиц красных волокон мышц. Тонус скелетных мышц связан с поступлением редких нервных импульсов к мышце, в результате чего мышечные волокна возбуждаются не одновременно, а попеременно. У домашних животных существуют специализированные рефлекторные дуги, одни из которых обеспечивают тетанические сокращения, а другие мышечный тонус. Тонус скелетных мышц играет важную роль в поддержании определенного положения тела в пространстве и деятельности двигательного аппарата.
Тетаническое сокращение мышц. Виды тетануса.
При воздействии на мышцу ритмических раздражений высокой частоты наступает сильное и длительное сокращение мышцы, которое называется тетаническим сокращением или тетанусом. Этот термин впервые применил Э. Вебер в 1821 году.
Тетанус может быть зубчатым (при частоте раздражений 20-40 Гц) или сплошным, гладким (при частоте 50 Гц и выше). Амплитуда тетанического сокращения в 2-4 раза выше амплитуды одиночного сокращения при той же силе раздражения.
Гладкий тетанус возникает тогда, когда очередной импульс раздражения действует на мышцу до начала фазы расслабления. При очень большой частоте раздражений каждое очередное раздражение будет попадать на фазу абсолютной рефрактерности и мышца вообще не будет сокращаться. Высота мышечного сокращения при тетанусе зависит от ритма раздражения, а также от возбудимости и лабильности, которые изменяются в процессе сокращения мышцы. Тетанус наиболее высокий при оптимальном ритме, когда каждый последующий импульс действует на мышцу в фазу экзальтации, вызванной предыдущим импульсом. В этом случае создаются наилучшие условия (оптимум силы и частоты раздражения, оптимум ритма) для работы мышцы.
При тетанических сокращениях мышечные волокна утомляются больше, чем при одиночных сокращениях. Поэтому даже в пределах одной мышцы происходит периодическая смена частоты импульсации (вплоть до полного исчезновения) в разных двигательных единицах.
Импульсы с мотонейронов в условиях покоя участвуют в поддержании мышечного тонуса.
Под тонусом понимают состояние естественного постоянного напряжения мышц при невысоких энергетических затратах. В поддержании тонуса участвуют проприорецепторы мышц (мышечные веретена) и центральная нервная система.
Осуществление тонуса скелетных мышц обусловлено функцией медленных двигательных единиц красных волокон мышц. Тонус скелетных мышц связан с поступлением редких нервных импульсов к мышце, в результате чего мышечные волокна возбуждаются не одновременно, а попеременно. У домашних животных существуют специализированные рефлекторные дуги, одни из которых обеспечивают тетанические сокращения, а другие мышечный тонус. Тонус скелетных мышц играет важную роль в поддержании определенного положения тела в пространстве и деятельности двигательного аппарата.
Тетанус, тетаническое мышечное сокращение— состояние длительного сокращения, непрерывного напряжения мышцы, возникающее при поступлении к ней через мотонейрон нервных импульсов с высокой частотой. При этом расслабления между последовательными одиночными сокращениями не происходит и возникает их суммация, приводящая к стойкому максимальному сокращению мышцы.
Различают зубчатый и гладкий тетанус. При зубчатом тетанусе каждый последующий нервный импульс воздействует на начавшую расслабляться мышцу, при этом происходит неполная суммация сокращений. При гладком тетанусе, имеющем большую амплитуду, воздействие импульса происходит в конце периода укорочения, что приводит к полной суммации сокращений.
13 Механизм мышечного сокращения (миофибриллы, саркомеры, сократительные белки).
Изменение механического состояния миофибриллярного сократительного аппарата мышечных волокон называется сокращением. Внешнее сокращение проявляется в изменении или напряжения, или длины мышцы, или и того, и другого. При этом потенциальная химическая энергия превращается в механическую и может совершаться механическая работа.
При произвольной внутренней команде сокращение мышцы человека начинается примерно через 0.05 с (50 м/с). За это время моторная команда передается от коры больших полушарий к мотонейронам спинного мозга и по двигательным волокнам к мышце. Подойдя к мышце, процесс возбуждения должен с помощью медиатора преодолеть нервно-мышечный синапс, что занимает примерно 0.5 м/с. Медиатором здесь является ацетилхолин, который содержится в синаптических пузырьках в пресинаптической части синапса. Нервный импульс вызывает перемещение синаптических пузырьков к пресинаптической мембране, их опорожнение и выход медиатора в синаптическую щель. Действие ацетилхолина на постсинаптическую мембрану чрезвычайно кратковременно, после чего он разрушается ацетилхолинэстеразой на уксусную кислоту и холин. По мере расходования запасы ацетилхолина постоянно пополняются путем его синтезирования в пресинаптической мембране. Однако, при очень частой и длительной импульсации мотонейрона расход ацетилхолина превышает его пополнение, а также снижается чувствительность постсинаптической мембраны к его действию, в результате чего нарушается проведение возбуждения через нервномышечный синапс. Эти процессы лежат в основе периферических механизмов утомления при длительной и тяжелой мышечной работе.
При достаточной частоте нервных импульсов ПКП достигает порогового значения и на мышечной мембране развивается мышечный потенциал действия. Он (со скоростью5 м/c)распространяется вдоль по поверхности мышечного волокна и заходит в поперечные трубочки внутрь волокна. Повышая проницаемость клеточных мембран, потенциал действия вызывает выход из цистерн и трубочек саркоплазматического ретикулума ионов Са2+, которые проникают в миофибриллы, к центрам связывания этих ионов на молекулах актина.
Под влиянием Са2+длинные молекулы тропомиозина проворачиваются вдоль оси и скрываются в желобки между сферическими молекулами актина, открывая участки прикрепления головок миозина к актину. Тем самым между актином и миозином образуются так называемые поперечные мостики. При этом головки миозина совершают гребковые движения, обеспечивая скольжение нитей актина вдоль нитей миозина с обоих концов саркомера к его центру, т. е, механическую реакцию мышечного волокна
14 Сила, работа и утомление мышц.
Основными показателями, характеризующими деятельность мышц, являются их сила и работоспособность.
Одиночное мышечное волокно развивает напряжение в 100-200 кг-сил во время сокращения.
Степень укорочения мышцы при сокращении зависит от силы раздражителя, морфологических свойств и физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие.
Незначительное растяжение мышцы, когда напрягаются упругие компоненты, является дополнительным раздражителем, увеличивает сокращение мышцы, а при сильном растяжении сила сокращения мышцы уменьшается.
Работа мышц. При изометрическом и изотоническом сокращении мышца совершает работу.
Оценивая деятельность мышц, обычно учитывают только производимую ими внешнюю работ)’.
Работа мышцы, при которой происходит перемещение груза и костей в суставах называется динамической.
Работа (W) может быть определена как произведение массы груза (Р) на высоту подъема (h)
Установлено, что величина работы зависит от величины нагрузки. Зависимость работы от величины нагрузки выражается законом средних нагрузок: наибольшая работа производится мышцей при умеренных (средних) нагрузках.
Максимальная работа мышцами выполняется и при среднем ритме сокращения (закон средних скоростей).
Мощность мышцы определяется как величина работы в единицу времени. Она достигает максимума у всех типов мышц так же при средних нагрузках и при среднем ритме сокращения. Наибольшая мощность у быстрых мышц.
При утомлении понижаются функциональные свойства мышцы: возбудимость, лабильность и сократимость. Высота сокращения мышцы при развитии утомления постепенно снижается. Это снижение может дойти до полного исчезновения сокращений. Понижаясь, сокращения делаются все более растянутыми, особенно за счет удлинения периода расслабления: по окончании сокращения мышца долго не возвращается к первоначальной длине, находясь в состоянии контрактуры (крайне замедленное расслабление мышцы). Скелетные мышцы утомляются раньше гладких. В скелетных мышцах сначала утомляются белые волокна, а потом красные.
15. Нейрон как структурно-функциональная единица ЦНС.
2. дендриты — короткие, сильноветвистые отростки, передающие информацию от периферии к телу нейрона.
Функции отростков заключаются в проведении информации к телу и от тела нейрона, в обеспечении взаимодействия нейронов с другими структурами.
По локализации нейроны подразделяются на центральные и периферические. Центральными называются те нейроны, тела которых лежат в пределах ЦНС. Периферические нейроны принадлежат периферической нервной системе. Они могут залегать в спинно-мозговых ганглиях, в ганглиях черепно- мозговых нервов, в ганглиях вегетативной нервной системы.
В зависимости от выполняемой функции нейроны делятся на 3 основные группы:
1. афферентные (чувствительные)
2. эфферентные (двигательные)
3. вставочные (контактные).
Это — специализированные клетки, способныепринимать,обрабатывать,кодировать,передавать ихранить информацию,реагировать на раздражения,устанавливать контакты с другими нейронами, клетками органов.
Уникальными особенностями нейрона являются способность генерировать электрические разряды и наличие специализированных окончаний — синапсов, служащих для передачи информации.
Число нейронов мозга человека приближается к 1011, на одном нейроне может быть 10000 синапсов, в каждом нейроне до 100000 нейротрубочек.
Если только эти элементы считать ячейками хранения информации, то нервная система может хранить 1019 единиц информации, что достаточно, чтобы вместить в ней практически все знания, накопленные человечеством. Поэтому вполне обосновано представление о способности человеческого мозга в течение жизни запоминать все, что происходит с организмом. Мозг, однако, не способен извлекать из памяти всю информацию, которая в нем хранится.
Функциональная Структура нейрона
Функционально нейрон состоит из следующих частей: воспринимающей — дендриты, мембрана сомы нейрона; интегративной — сома с аксонным холмиком; передающей —аксонный холмик с аксоном.
16. Функциональная классификация нейронов.
Функционально нейроны делят на три типа:
Афферентные — выполняют функцию получения и передачи информации в вышележащие структурыЦНС.
Промежуточные — обеспечивают взаимодействие между нейронами одной структуры.
Эфферентные — за счет длинного аксона передают информацию в нижележащие структуры ЦНС, в нервные узлы, лежащие за ее пределами, и в органы организма.
По форме нейроны делят на:
Биполярные, Униполярные, Псевдоуниполярные, Мультиполярные.
По химической характеристике выделяемых в окончаниях аксонов веществ, отличают нейроны:
Серотонинэргические и др.
По признаку чувствительность к разным раздражителям нейроны делят на
Моносенсорные нейроны располагаются чаще в первичных проекционных зонах коры и реагируют только на сигналы своей модальности. Например, значительная часть нейронов первичной зрительной коры реагирует только на световое раздражение сетчатки глаза.
Бисенсорные нейроны располагаются преимущественно во вторичных зонах коры анализатора и могут реагировать как на сигналы своей, так и на сигналы другой модальности. Например, нейроны вторичной зрительной коры реаги-руют на зрительные и слуховые раздражения.
Полисенсорные нейроны — это чаще всего нейроны ассоциативных зон мозга. Они способны реагировать на раздражение слуховой, зрительной, кожной и др. анализаторных систем.
Тетанус
Тетаническое сокращение
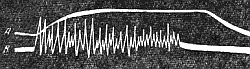 | Рис. 142. Одновременная запись тетанического сокращения (А) и токов действия (В) мышцы (по А. Г. Гинецинскому). |
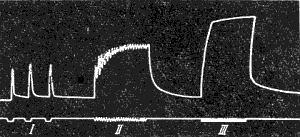
Тетанические сокращения мышцы представля ют собой результат суммации одиночных сокращений. Для исследования суммации сокращений наносят на мышцу два одиночных раздражения. Промежуток времени между раздражениями должен быть такой, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого сокращения. При этом возможны два случая.
Если второе раздражение поступает, когда мышца уже начала расслабляться после первого сокращения, то на миографической кривой вершина второго сокращения будет отделена от вершины первого небольшим западением кривой ( рис. 143, Б ). Если же второе раздражение действует, когда первое сокращение ещё не дошло до своей вершины, то второе сокращение полностью сливается с первым, образуя вместе с ним единую суммированную вершину ( рис. 143, А ).
 |
 |
 |
| Медленные двигательные единицы | Быстрые двигательные мышцы |
| Состоят из красных мышц | Состоят из белых мышечных волокон |
| Содержат меньше мышечных волокон, но каждое мышечное волокно содержит много миоглобина и МХ | В них мало митохондрий, но много гликолитических ферментов. Вследствие этого анаэробный распад гликогена в них происходит с большой скоростью. Однако поступление в П-ПМ глюкозы из крови идет медленно. И медленно утилизируется |
| Мышечные волокна обладают высокой способностью к аэробному окислению глюкозы, ЖК, кетоновых тел | Быстрее переходят от состояния покоя к максимальной активности |
| Хорошо кровоснабжаются | Сокращаются энергично, но скоро наступает утомление, так как запасы гликогена в белых мышечных волокнах быстрее истощаются |
| Развивают меньшую силу при сокращении | Являются высокопороговыми, быстрыми и сильными. Вовлекаются в возбуждение, когда необходимо осуществить сильное напряжение. |
| – могут работать без утомления дольше | |
| – мотонейроны являются более низкопороговыми, то есть вовлекаются в возбуждение раньше | |
| – более приспособлены к продолжительной работе | |
| Активация обеспечивает поддержание позы (иначе называются позотоническими) |
В теле человека нет целиком былых или только красных мышц в отличие от многих животных (кроликов или птиц).
Мышцы человека содержат и красные, и белые мышечные волокна, относительное количество которых в разных мышцах неодинаковое.
Имеются и индивидуальные различия. Например, более перспективным спринтером можно считать человека в мышцах которого много белых мышечных волокон.
Быстрые и медленные мышечные единицы теплокровных животных относят к фазным единицам
У амфибий, рептилий, в некоторых мышцах теплокровных (например, наружные мышцы глаза) содержатся тонические двигательные единицы. Они не подчиняются закону «все или ничего». Возбуждение носит характер локального ответа. Их сокращение осуществляется медленнее, чем сокращение фазных единиц.
Механизм мышечного сокращения
Клетки мышечной ткани содержат специфичные субклеточные структуры – миофибриллы, состоящие из нитей – филаментов, собранных в пучки и располагающихся вдоль клетки.
В П-ПМ ткани миофибриллы строго упорядочены, образуют чередующиеся темные и светлые диски, обладающие разными оптическими свойствами.
Темные диски А образованы толстыми нитями, основным белком которых является миозин. Светлые диски I образованы тонкими нитями – из белка актина.
Актин и миозин – это сократительные фибриллярные белки, главный компонент всех сократительных систем.
Каждая миофибрилла разделена Z-линиями на сакромеры. Саркомер – комплекс из 1-го темного диска и 2-х прилежащих к нему половин светлых дисков. Многие сотни саркомеров образуют миофибриллы.

Миозиновые нити имеют поперечные мостики (выступы) с головками, которые отходят от нити миозина биполярно.
Актиновая нить состоит из 2-х, закрученных одна вокруг другой цепочек молекул актина. На нитях актина расположены молекулы тропонина, а в желобках, между двумя нитями актина лежат нити тропомиозина. В состоянии покоя молекулы тропомиозина расположены так, что они недоступны для взаимодействия и прикрепления поперечных мостиков миозина к актиновым центрам.
Сокращение мышц есть результат укорочения каждого саркомера. Укорочение которого происходит путем сдвигания нитей актина между миозиновыми нитями. При сокращении саркомер укорачивается на 25-50%, при этом диски I укорачиваются, а диски А сохраняют свой размер. Нити миозина и актина связаны между собой поперечными мостиками.
И это своеобразный механизм сцепления этих белковых молекул. При сокращении мышцы, нити актина скользят по нитям миозина при помощи мостиков, вдвигаясь между миозиновыми нитями.
Ионизированный кальций активирует активные центры молекул миозина, представленные кальциевой АТФазой. Так как этот фермент активируется, происходит расщепление молекулы АТФ, вследствие чего выделяется энергия, необходимая для сокращения мышечных волокон.
Ca 2+ вызывают конформационные изменения в системе белков тропонин-тропомиозин-актин: кальций связывает белок тропонин, который делает недоступным активный центр молекулы актина для миозина. Следовательно на нитях актина открываются активные центры для связывания с активными центрами миозина. Вследствие этого между этими молекулами формируются поперечные мостики (химические связи) – механизм сцепления молекул сократительных белков. Вследствие этого нити актина вдвигаются между нитями миозина.
Концентрация свободных ионов Ca 2+ в цитоплазме П-ПМ волокна низкая и составляется примерно 1мкМоль на 1 гр. Депо кальция в мышечной клетке является саркоплазматический ретикулум (СПР). Количество кальция в сакроплазматическом ретикулуме примерно равно концентрации кальция во внеклеточной жидкости. Выход Ca 2+ из СПР в саркоплазму осуществляется за счет системы поперечных трубочек (Т-система) в сарколемме, которые представляют собой впячивание (инвагинацию) поверхностной мембраны, то есть сарколеммы.
По обе стороны поперечных трубочек расположены цистерны. Мембрана поперечных трубочек по своим электро-физиологическим свойствам сходна с поверхностной мембраной и способна к генерации потенциала действия. При возбуждении мышечного волокна происходит деполяризация мембраны и потенциал действия распространяется по сарколемме на мембрану поперечных трубочек. Вследствие этого происходит деполяризация –> натрий устремляется в клетку –> концентрация натрия в саркоплазме возрастает –> происходит открывание кальциевых ионных каналов мембран СПР –> ионы кальция из СПР устремляются в саркоплазму клетки –> концентрация кальция увеличивается –> запускается механизм сократительных белков актина и миозина, так как активные центры этих белков становятся доступными для взаимодействия и формирования поперечных мостиков – происходит сокращения мышечного волокна.
Таким образом Ca 2+ обеспечивают сопряжение двух процессов: возбуждения и сокращения.
Взаимодействие актина и миозина активирует АТФазу миозина – АТФ расщепляется и выделяется энергия, которая обеспечивает разъединение молекул актина и миозина.
Алгоритм сократительного акта. Последовательность событий (алгоритм), ведущих к сокращению и последующему расслаблению П-ПМ волокон может быть представлен следующей схемой:
2) Вследствие этого проведение потенциала действия вдоль сарколеммы и вглубь мышечного волокна по Т-системе трубочек.
3) Вследствие этого выход Ca 2+ из цистерн СПР.
4) Увеличение концентрации Ca 2+ в саркоплазме.
5) Диффузия Ca 2+ к миофибриллам.
6) Взаимодействие Ca 2+ с тропонином.
7) Активация Ca 2+ кальциевой АТФазы миозина.
8) Тормозящее влияние тропонина на активный центр актина устраняется.
9) Между активными центрами молекул актина и миозина формируются поперечные мостики.
10) Нити актина скользят вдоль миозиновых нитей.
11) Происходит сокращение мышечного волокна. Следовательно повышается активность кальциевой АТФазы миозина
12) Молекула АТФ расщепляется, выделяется энергия, которая используется для разъединения, следовательно происходит расслабление мышцы.
Таким образом для того, чтобы процесс связывания актина и миозина носил обратимый характер необходимо присутствие АТФ, если концентрация АТФ падает ниже некоторой критической величины, тогда комплекс актин+миозин становится стабильным.



