Как устроен продолговатый мозг?
Центральная нервная система разделена на отделы, которые передают друг другу информацию посредством импульсов. Ее работа контролируется механизмом из структур, объединенных нервными волокнами. Продолговатый мозг является продолжением спинного мозга.

Структура

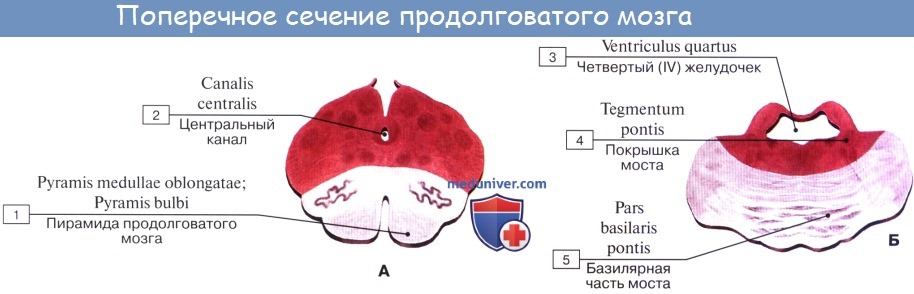
Продолговатый мозг располагается на пересечении головного и спинного мозга. В районе затылочного отверстия спинной мозг немного толще и похож на луковицу, благодаря чему он получил такое второе название. Луковица объединяет малый мозг и мост. Продолговатый отдел имеет срединное углубление, которое является продолжением борозды спинного мозга. Рядом с углублением расположены пирамиды, переходящие в спинномозговые канатики. Дорсальную часть борозды можно обнаружить в задней части структуры. По бокам располагаются канатики, направляющиеся к мозжечку.
Структуру луковицы нельзя назвать однородной. Нейроны окружены ядрами, а длинные волокна идут через спинной мозг, а затем от них отходят небольшие отростки. Ядра серого вещества объединяются при помощи данных отростков, благодаря чему происходит функционирование органа. Сама же луковица имеет внутреннюю и наружную структуры.
Наружное строение
Внешняя часть луковицы образована из парных долей, которые имеют конусообразную форму. По бокам от пирамид находится олива, в которой содержатся ядра. Задняя часть луковицы имеет разделительную борозду. Тыльная сторона продолговатого отдела имеет связки и располагается около ромбовидной нижней части. Пирамидная система выполняет функцию координации движений.
Внутреннее строение
Внутреннее строение представлено в виде ядер. В разрезе продолговатые структуры имеют борозды и оливы. Какова роль ядра в оливе? Оно контролирует умение человека принимать вертикальное положение. Около пирамидного тракта и оливы есть углубления. Связь с другими отделами происходит посредством передачи импульсов.
Ядра черепных нервов
Нервные центры отвечают за функциональность структуры. Ядра черепных нервов представлены управляющими центрами. Они имеют такой состав:
Функциональные особенности
С рождения продолговатый отдел отвечает только за основные функции: дыхание, кровообращение и пищеварение. Полноценно данная структура начинает работать лишь с 7-летнего возраста. К основным функциям продолговатой структуры относятся:
Все рефлексы создаются в продолговатом мозге, а следующим шагом является передача импульса в заднюю мозговую структуру. Функции продолговатого мозга разделяются на жизненно важные и рефлекторные. Также можно поделить возможности луковицы на:
Надо отметить, что мозг нуждается в постоянных тренировках. Иначе постепенно его эффективность снижается. Улучшайте работу мозга с помощью тренажеров Викиум!
Что делает продолговатый мозг у млекопитающих
Medulla oblongata имеет вид луковицы, bulbus cerebri (отсюда термин «бульбарные расстройства»); верхний расширенный конец граничит с мостом, а нижней границей служит место выхода корешков I пары шейных нервов или уровень большого отверстия затылочной кости.
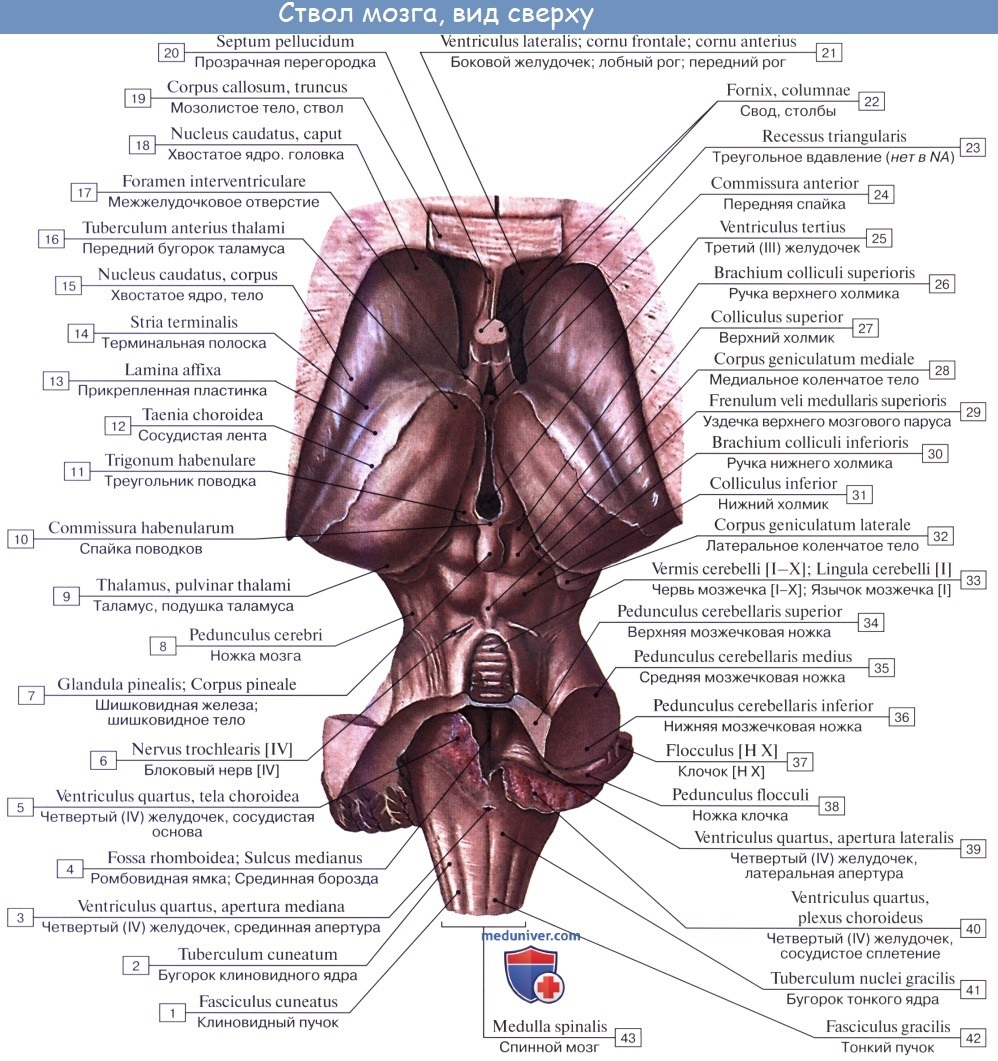
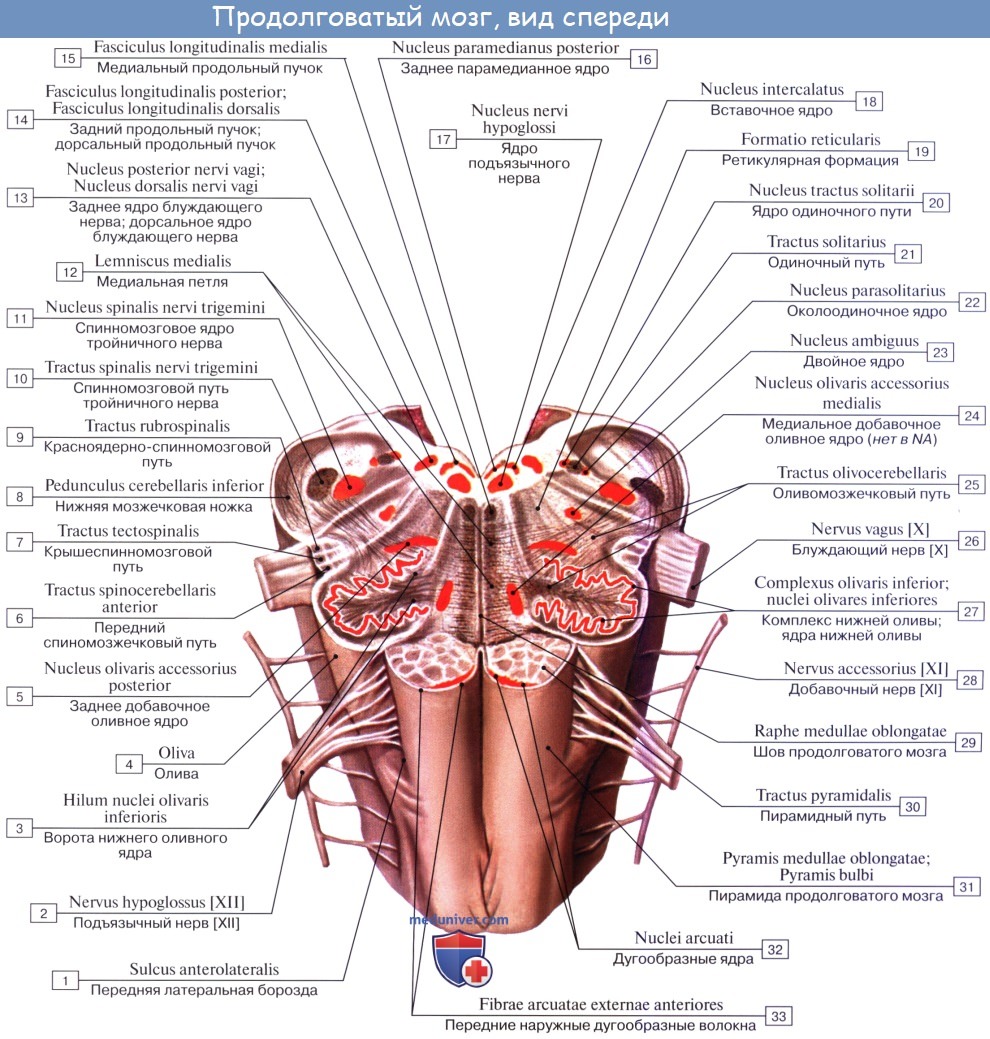
1. На передней (вентральной) поверхности продолговатого мозга по средней линии проходит fissura mediana anterior, составляющая продолжение одноименной борозды спинного мозга. По бокам ее на той и другой стороне находятся два продольных тяжа — пирамиды, pyramides medullae oblongatae, которые как бы продолжаются в передние канатики спинного мозга.
Составляющие пирамиды пучки нервных волокон частью перекрещиваются в глубине fissura mediana anterior с аналогичными волокнами противоположной стороны — decussatio pyramidum, после чего спускаются в боковом канатике на другой стороне спинного мозга — tractus corticospinal (pyramidalis) lateralis, частью остаются неперекрещенными и спускаются в переднем канатике спинного мозга на своей стороне — tractus corticospinalis (pyramidalis) anterior.
Пирамиды отсутствуют у низших позвоночных и появляются по мере развития новой коры; поэтому они наиболее развиты у человека, так как пирамидные волокна соединяют кору большого мозга, достигшую у человека наивысшего развития, с ядрами черепных нервов и передними рогами спинного мозга,
Латерально от пирамиды лежит овальное возвышение — оливa, oliva, которая отделена от пирамиды бороздкой, sulcus anterolateral.
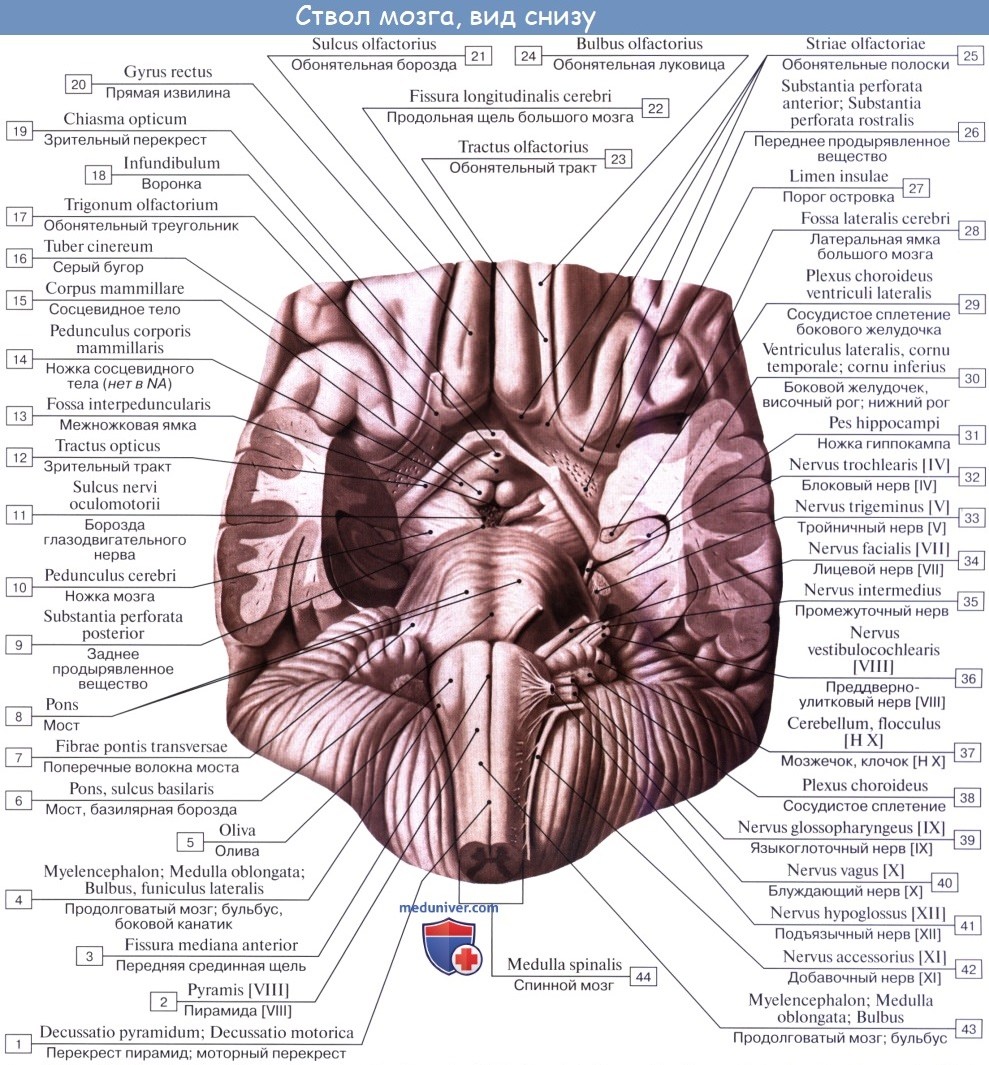
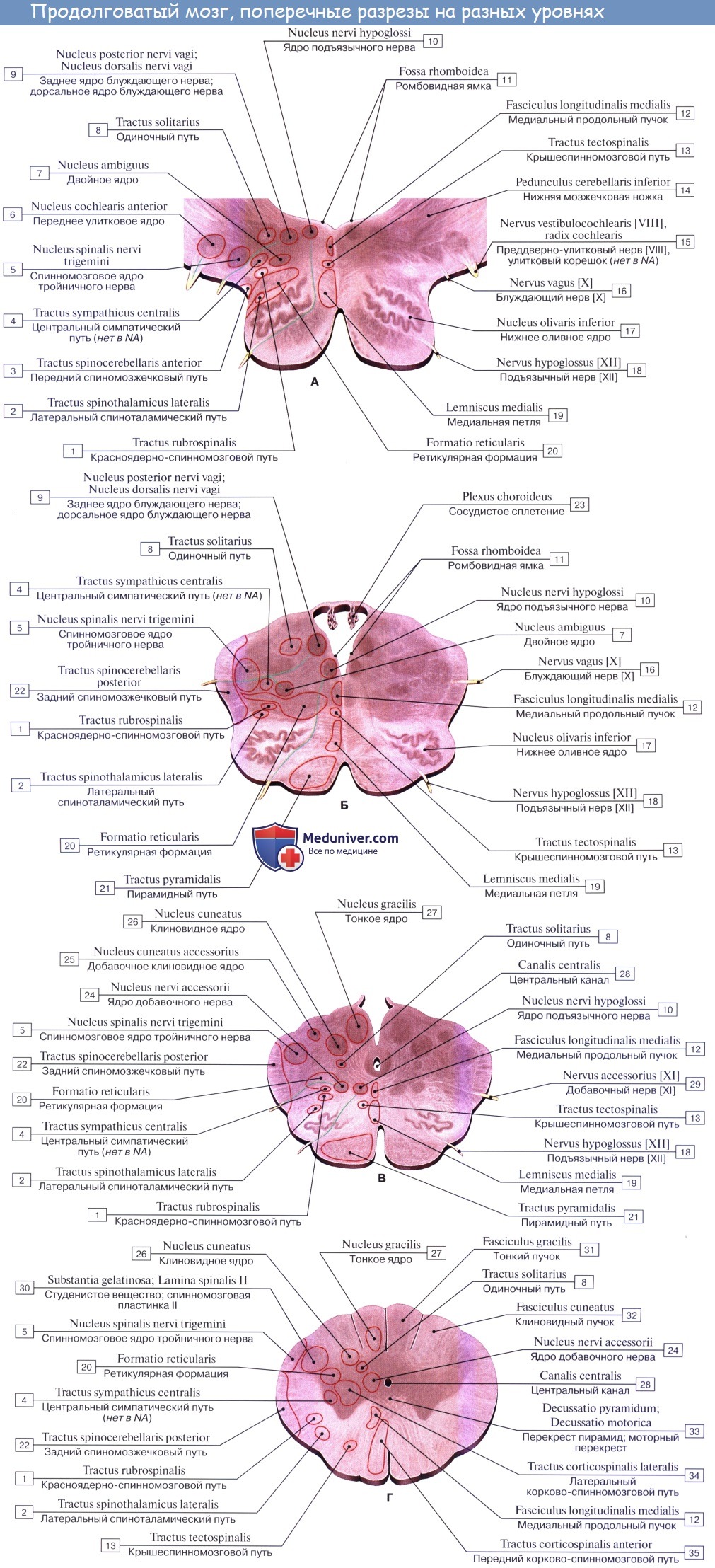
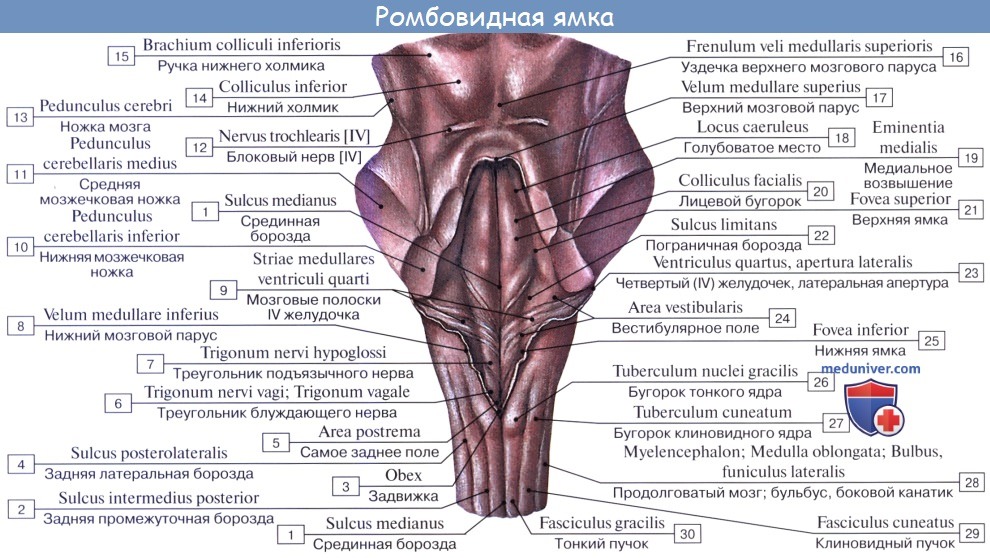
2. На задней (дорсальной) поверхности продолговатого мозга тянется sulcus medianus posterior — непосредственное продолжение одноименной борозды спинного мозга. По бокам ее лежат задние канатики, ограниченные латерально с той и другой стороны слабо выраженной sulcus posterolaterals. По направлению кверху задние канатики расходятся в стороны и идут к мозжечку, входя в состав его нижних ножек, pedunculi cerebellares inferiores, окаймляющих снизу ромбовидную ямку.
Каждый задний канатик подразделяется при помощи промежуточной борозды на медиальный, fasciculus gracilis, и латеральный, fasciculus cuneatus. У нижнего угла ромбовидной ямки тонкий и клиновидный пучки приобретают утолщения — tuberculum gracilum и tuberculum cuneatum. Эти утолщения обусловлены соименными с пучками ядрами серого вещества, nucleus gracilis и nucleus cuneatus.
В названных ядрах оканчиваются проходящие в задних канатиках восходящие волокна спинного мозга (тонкий и клиновидный пучки). Латеральная поверхность продолговатого мозга, находящаяся между sulci posterolateralis et anterolateralis, соответствует боковому канатику. Из sulcus posterolateralis позади оливы выходят XI, X и IX пары черепных нервов. В состав продолговатого мозга входит нижняя часть ромбовидной ямки.
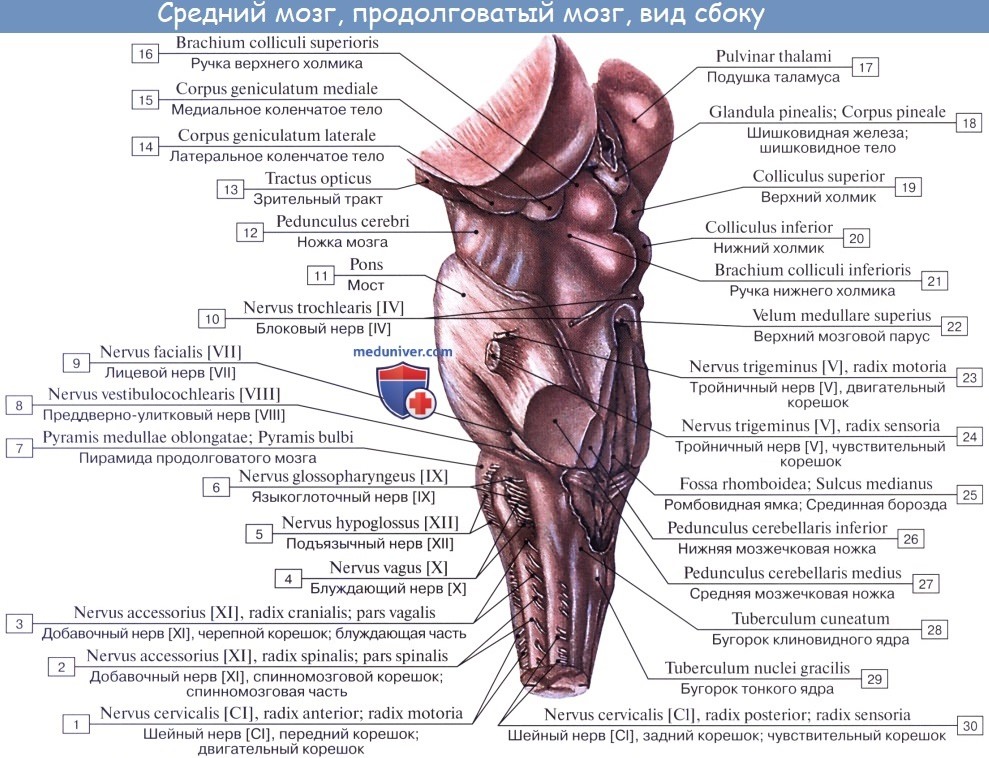
Внутреннее строение продолговатого мозга. Продолговатый мозг возник в связи с развитием органов гравитации и слуха, а также в связи с жаберным аппаратом, имеющим отношение к дыханию и кровообращению. Поэтому в нем заложены ядра серого вещества, имеющие отношение к равновесию, координации движений, а также к регуляции обмена веществ, дыхания и кровообращения.
1. Nucleus olivaris, ядро оливы, имеет вид извитой пластинки серого вещества, открытой медиально (hilus), и обусловливает снаружи выпячивание оливы. Оно связано с зубчатым ядром мозжечка и является промежуточным ядром равновесия, наиболее выраженным у человека, вертикальное положение которого нуждается в совершенном аппарате гравитации. (Встречается еще nucleus olivaris accessorius medialis.)
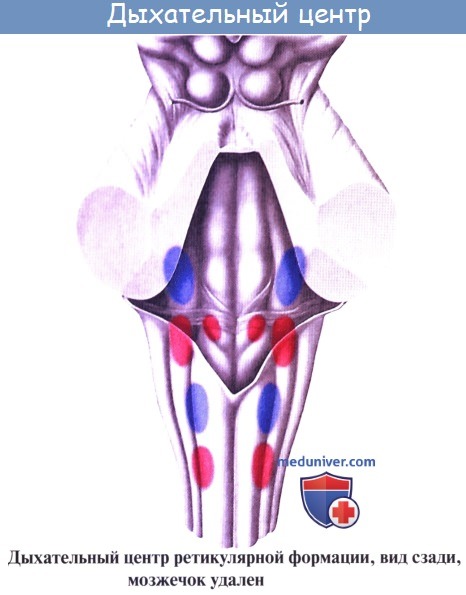
2. Formatio reticularis, ретикулярная формация, образующаяся из переплетения нервных волокон и лежащих между ними нервных клеток.
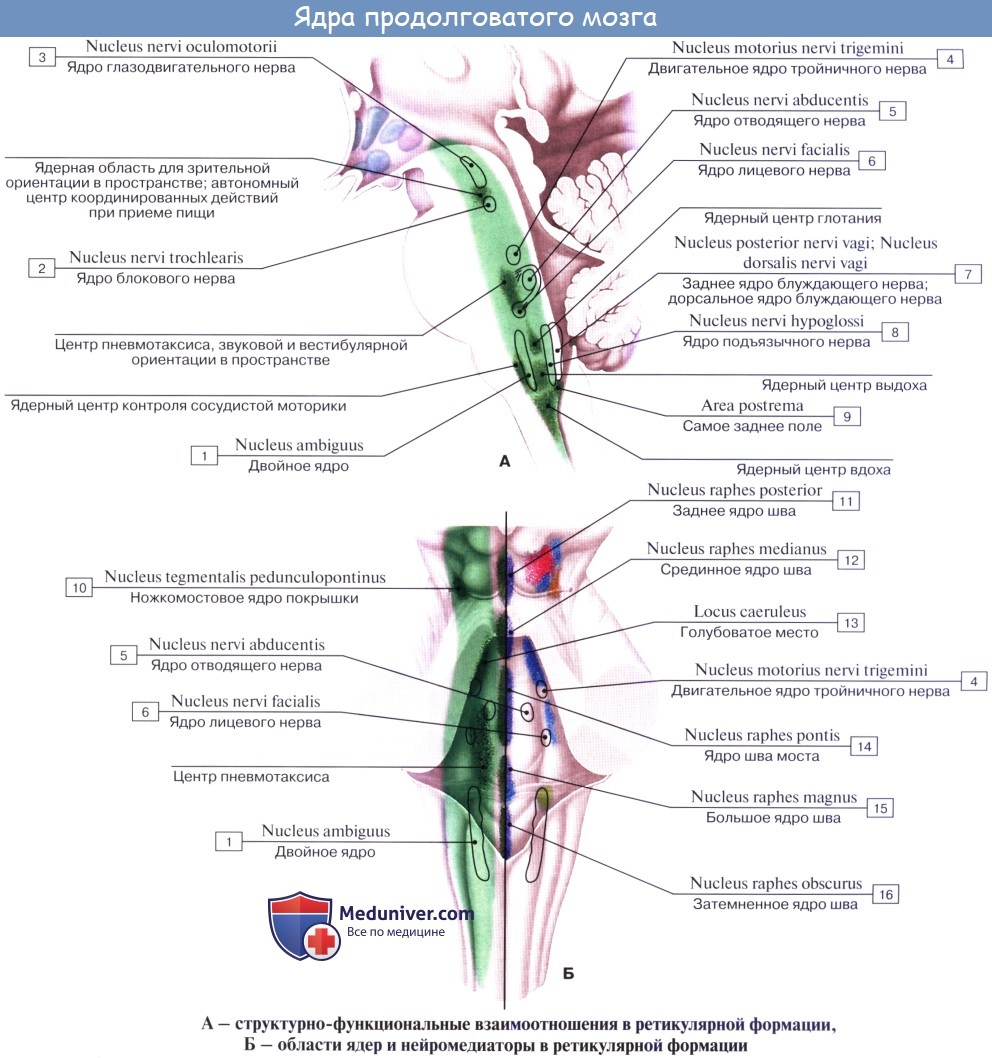
3. Ядра четырех пар нижних черепных нервов (XII —IX), имеющие отношение к иннервации производных жаберного аппарата и внутренностей.
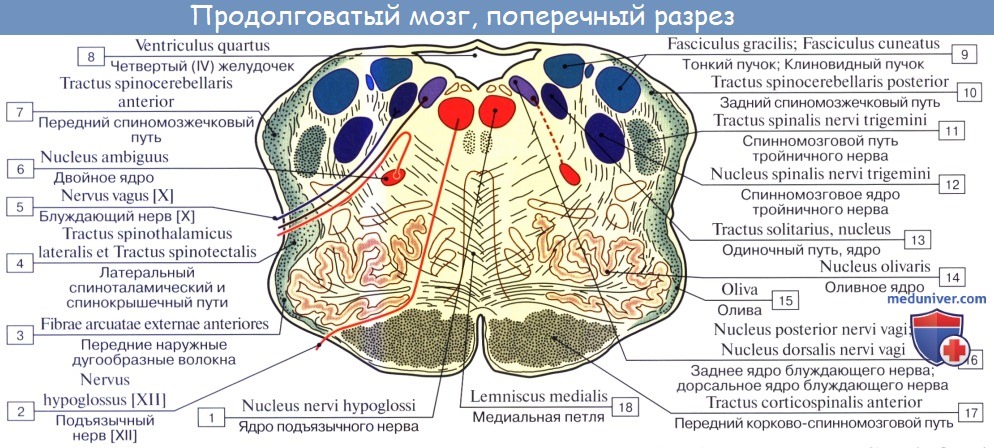
Белое вещество продолговатого мозга содержит длинные и короткие волокна. К длинным относятся проходящие транзитно в передние канатики спинного мозга нисходящие пирамидные пути, частью перекрещивающиеся в области пирамид. Кроме того, в ядрах задних канатиков (nuclei gracilis et cuneatus) находятся тела вторых нейронов восходящих чувствительных путей. Их отростки идут от продолговатого мозга к таламусу, tractus bulbothalamicus.
Волокна этого пучка образуют медиальную петлю, lemniscus medialis, которая в продолговатом мозге совершает перекрест, decussatio lemniscorum, и в виде пучка волокон, расположенных дорсальнее пирамид, между оливами — межоливныи петлевой слой — идет далее. Таким образом, в продолговатом мозге имеется два перекрестка длинных проводящих путей: вентральный двигательный, decussatio pyramidum, и дорсальный чувствительный, decussatio lemniscorum.
К коротким путям относятся пучки нервных волокон, соединяющие между собой отдельные ядра серого вещества, а также ядра продолговатого мозга с соседними отделами головного мозга. Среди них следует отметить tractus olivocerebellaris и лежащий дорсально от межоливного слоя fasciculus longitudinalis medialis.
Топографические взаимоотношения главнейших образований продолговатого мозга видны на поперечном срезе, проведенном на уровне олив. Отходящие от ядер подъязычного и блуждающего нервов корешки делят продолговатый мозг на той и другой стороне на три области: заднюю, боковую и переднюю. В задней лежат ядра заднего канатика и нижние ножки мозжечка, в боковой — ядро оливы и formatio reticularis и в передней — пирамиды.
Каковы особенности строения головного мозга млекопитающих? Анатомия млекопитающих
Головной мозг — это один из органов, составляющих центральную нервную систему млекопитающих. Он служит основным регулятором работы организма. Для понимания его функционирования важно ответить на вопрос о том, каковы особенности строения головного мозга у млекопитающих. А также имеет смысл проследить этапы его эволюционного развития.
Общая характеристика головного мозга млекопитающих
Мозг млекопитающих состоит из тех же отделов, что и мозг остальных позвоночных. Каковы особенности строения головного мозга млекопитающих в сравнении с другими позвоночными? Для млекопитающих характерен большой объем и сложнейшее устройство полушарий переднего мозга (прозэнцефалона). Нередко встречается ошибочное мнение о том, что кора головного мозга впервые появилась у млекопитающих. Это не так — впервые она появилась у пресмыкающихся, а вот интенсивно развиваться начала лишь у млекопитающих. У млекопитающих, обладающих высоким уровнем организации, кора образует множество борозд и извилин. За счет этого ее площадь значительно увеличена. Также существенное развитие получили средний мозг и мозжечок головного мозга. Это объясняется с тем, что для млекопитающих характерны высокая моторная активность, сложная система как рефлексов, так и органов чувств.
Эволюция головного мозга млекопитающих
Общее направление в эволюционном развитии мозга позвоночных — увеличение объема, особенно переднего мозга, и специализация высших мозговых центров, согласовывающих деятельность органов, сложного поведения и функционирование всего организма. Мозг млекопитающих крупный за счет увеличения коры, а именно складчатости его плаща. Появились структуры, обеспечивающие взаимосвязи коры и других отделов ЦНС. Для связи мозжечка и больших полушарий в заднем мозге возникает варолиев мост. Появившаяся новая кора берет на себя выполнение функций высших сенсорных систем. У высших млекопитающих большая часть коры — ассоциативные зоны, выполняющие интегративную функцию. У приматов очень развиты извилины коры полушарий мозга. В извилинах локализованы двигательные, ассоциативные, чувствительные центры, регулирующие высшую нервную деятельность. Для приматов характерно сложнейшее поведение. Homo sapiens, относящийся к отряду приматов, имеет развитые большие полушария, где расположены центры речи и второй сигнальной системы, связанной с сознанием и абстрактным мышлением.
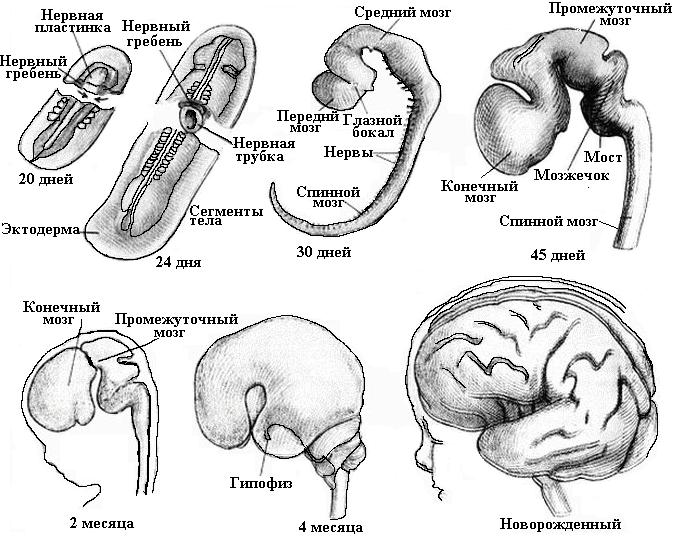
Строение головного мозга
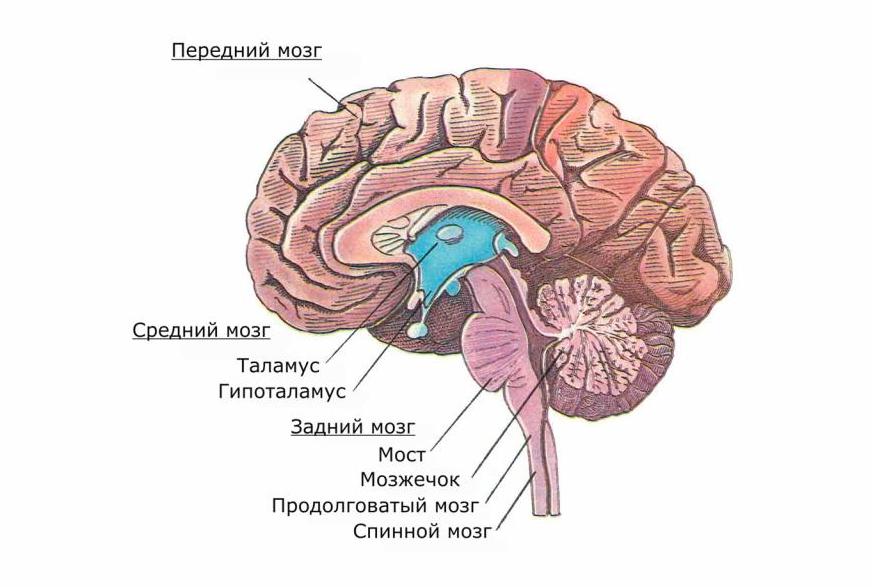
Продолговатый мозг
В полости черепа спинной мозг без особого морфологического изменения переходит в продолговатый мозг. В противоположность спинному мозгу он не имеет сегментированного и повторяемого строения. В продолговатом мозге серое вещество располагается в переключательных черепно-мозговых ядрах и в ретикулярной формации. Ретикулярной формацией (или сетчатым веществом) называется группа нервных клеток с отростками, расположенных по всей длине спинного, продолговатого и промежуточного мозга.
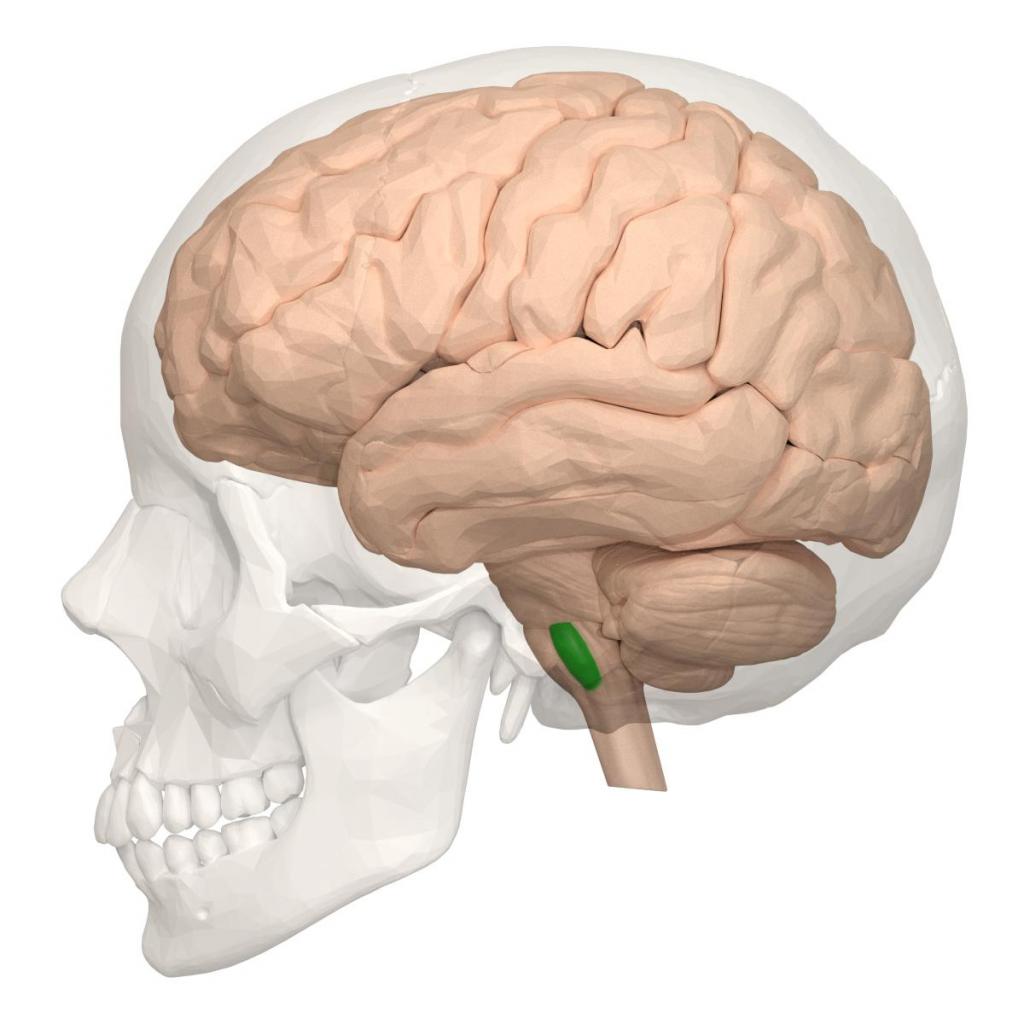
Варолиев мост
Содержит ядра черепно-мозговых нейронов, образованных из серого вещества. Проводящие пути варолиева моста соединяют продолговатый мозг и мозжечок с большими полушариями.
Функции варолиева моста:
Средний мозг
Включает две ножки и крышу и связывает задний и промежуточный мозг. На его крыше (пластинке четверохолмия) находятся:
Выполняет важную функцию регуляции тонуса мышц, сохранения пространственного положения тела и является центром ориентировочных рефлексов.
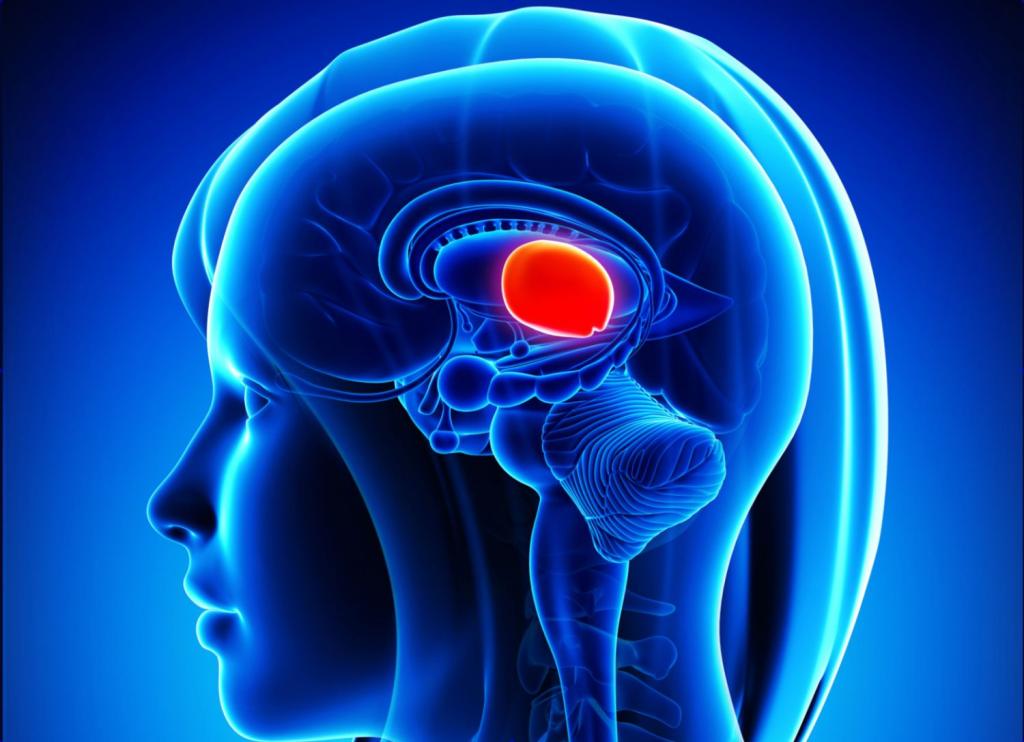
Передний мозг
Включает промежуточный мозг и конечный, состоящий из больших полушарий мозга.
Промежуточный — состоит из гипоталамуса, таламуса, метаталамуса и эпиталамуса. Передний мозг — это наиболее развитый отдел головного мозга у млекопитающих
Метаталамусом называют подкорковый центр слуха и зрения, а эпиталамусом — надбугорную часть промежуточного мозга.
В эпиталамус входит эпифиз (другое название — шишковидная железа), выполняющий эндокринные функции.
Таламус (зрительные бугры) — подкорковый центр различных видов чувствительности, кроме обоняния. В таламусе осуществляется первичная обработка информации об ощущениях, например, впервые осознается изменение температуры, зрительных, слуховых образов и т.д.
Гипоталамус образован скоплениями нейронов, имеющих следующие особенности:
Функции гипоталамуса следующие:
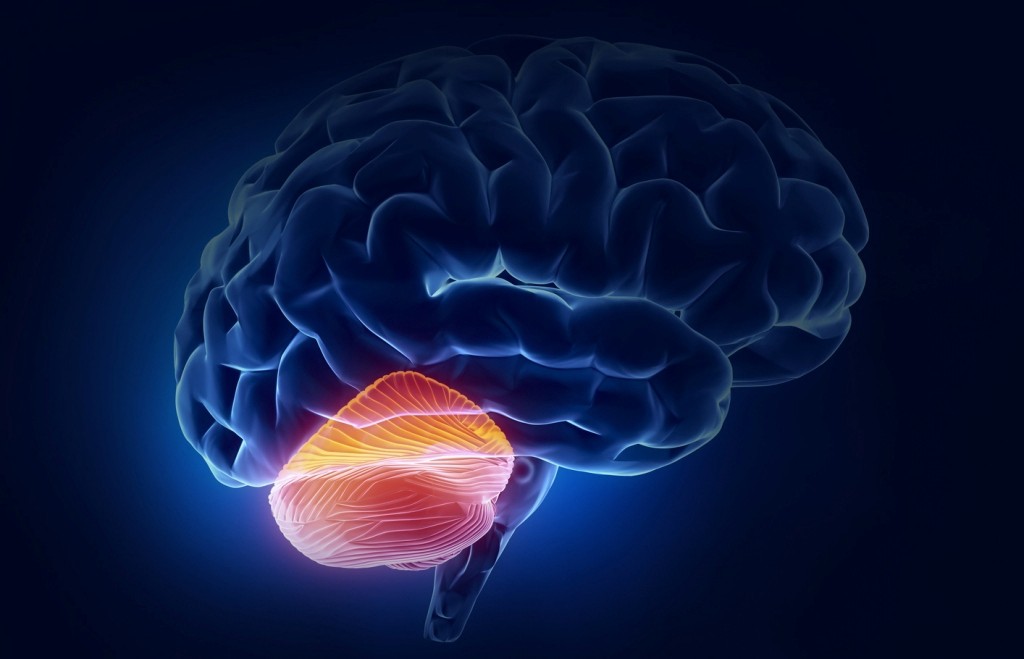
Мозжечок
Конечный мозг
Включает два полушария, занимает примерно 80% массы всего головного мозга, покрывая сверху другие отделы. Полушария являются самыми развитыми отделами мозга млекопитающих. Большие полушария головного мозга у млекопитающих разделяются щелью. В ее углублении находится мозолистое тело, соединяющее их.
В состав больших полушарий входит серое вещество, из которого образуются кора полушарий, подкорковые ядра, а также белое вещество, формирующее проводящие нервные пути.
Полушария делятся тремя бороздами, центральной, боковой и затылочно-теменной, на четыре доли: теменную, затылочную, лобную и височную.
Исходя из выполняемых функций, кора головного мозга разделена на три зоны:
Масса мозга
Уникальность строения мозга человека
Каковы особенности строения головного мозга млекопитающих, мы в общих чертах выяснили. Чем же отличается от прочих мозг гоминоидов, к которым относится человек?
Исследователям всегда было интересно, с чем связано то, что мозг человека в ходе развития формирует новые возможности. Как выяснилось, он с точки зрения анатомии не имеет исключительно человеческих структур. Таким образом, новая функция возникает, но структуры, ответственной за нее, нет. Уникальность заключается в том, что человеческий мозг очень пластичен. Пластичность проявляется на двух уровнях. Первый уровень состоит в восприимчивости к новой информации и отсутствии предопределенности поведения. Второй — в пластичности нейронов головного мозга. Пример подобной пластичности — реабилитация после инсульта с восстановлением способностей.
При изучении форм черепов исследователями было обнаружена интересная закономерность: черепа взрослых людей схожи с черепами детенышей человекообразных обезьян, таких как шимпанзе и гориллы. После этого наблюдения палеонтолог, биолог-эволюционист Стивен Гулд высказал предположение о том, что, возможно, причина заключается не в костях, а в том, что мозг человека даже в зрелом возрасте не утрачивает инфантильных характеристик. Таким образом сохраняется поведенческая гибкость, свойственная обезьянам юного возраста, которую они утрачивают, взрослея. Поэтому человек может обучаться новому в течение всей жизни. Ему многое интересно и он имеет больше социальных связей.
Рост мозга у древних млекопитающих был связан с развитием обоняния

Американские палеонтологи при помощи компьютерной рентгеновской томографии изучили эндокасты (мозговые полости) двух раннеюрских маммалиаформ — животных, переходных между зверозубыми рептилиями цинодонтами и первыми млекопитающими. Исследование показало, что становление млекопитающих сопровождалось значительным увеличением мозга, которое проходило в три этапа. На первом этапе увеличились отделы мозга, отвечающие за обоняние и сенсомоторные функции (осязание и координацию движений); два последующих этапа отражают дальнейшее совершенствование обоняния.
Эволюционная история наземных позвоночных (тетрапод) началась с того, что в конце девонского периода (380–360 млн лет назад) одна из групп древних лопастепёрых рыб дала начало первым амфибиям. В следующем, каменноугольном периоде от амфибий произошли рептилии, которые вскоре подразделились на несколько эволюционных линий. Важнейшую роль в последующей истории наземной фауны сыграли две из них: диапсиды (см. также Diapsid) и синапсиды (см. также Synapsid). Диапсидные рептилии подразделилась на архозавров (к которым относятся, в частности, динозавры и произошедшие от них птицы) и лепидозавров (ящерицы, змеи и другие). Синапсидные рептилии были многочисленны и разнообразны в пермском и триасовом периодах, но потом мало-помалу вымерли, за исключением одной группы, которая дала начало млекопитающим.
Эволюционный переход от синапсидных рептилий к млекопитающим был длительным и постепенным; изучен он весьма подробно (см.: маммализация териодонтов). Древнейшие ископаемые находки животных, которых палеонтологи безоговорочно считают «настоящими млекопитающими», имеют позднетриасовый возраст (немногим более 200 млн лет назад). Непосредственных предков первых млекопитающих относят к группе «маммалиаформ», которые, в свою очередь, представляют собой одну из ветвей цинодонтов. Цинодонты являются одной из групп териодонтов, или зверозубых ящеров, а териодонты — это одна из эволюционных ветвей синапсид.
Палеонтологи детально реконструировали основные этапы эволюционных преобразований зубов и скелета териодонтов по мере их «маммализации» — постепенного эволюционного движения в сторону млекопитающих. Об эволюции мозга известно значительно меньше. Между тем очевидно, что именно прогрессивное развитие мозга во многом предопределило эволюционный успех млекопитающих.
Мозг млекопитающих радикально отличается от мозга рептилий, в том числе цинодонтов, не только своим размером, но и строением. В частности, у млекопитающих развилась так называемая «новая кора» — неокортекс (см. также Neocortex), отвечающая за сенсомоторные функции, резко увеличились обонятельные луковицы и отделы коры, связанные с обонянием, а также мозжечок. Но о том, когда и в какой последовательности произошли эти изменения в ходе эволюционного становления млекопитающих, до сих пор было известно очень мало.

Изучение мозга маммалиаформ и первых млекопитающих затруднялось, во-первых, редкостью находок хорошо сохранившихся черепов, во-вторых — тем обстоятельством, что для изучения эндокаста (слепка мозговой полости, по которому можно судить о размере и форме мозга) череп, как правило, нужно было разрушить.
Статья американских палеонтологов, опубликованная в последнем номере журнала Science, в значительной мере заполняет этот досадный пробел. При помощи компьютерной рентгеновской томографии авторам удалось, не разрушая драгоценных черепов, получить детальные трехмерные изображения эндокастов двух маммалиаформ, живших в начале ранней юры (200–190 млн лет назад) на территории нынешнего Китая.
Изученные маммалиаформы Morganucodon oehleri и Hadrocodium wui — ближайшие родственники первых «настоящих» млекопитающих. По строению скелета они представляют собой классические переходные формы между «еще рептилиями» и «уже млекопитающими». При этом Morganucodon стоит ближе к «базальным» (примитивным) цинодонтам, а Hadrocodium настолько близко подошел к млекопитающим, насколько это возможно, оставаясь формально за пределами группы. Исследование показало, что по строению мозга эти животные тоже занимают промежуточное положение между типичными цинодонтами и их потомками — млекопитающими.
Ранее были изучены эндокасты базальных цинодонтов Thrinaxodon и Diademodon. Выяснилось, что мозг у них был еще вполне «рептильный» по своему размеру и строению.



Мозг Morganucodon, как выяснилось, был гораздо больше похож на мозг млекопитающего. По объему он в полтора раза превосходит мозг базальных цинодонтов (EQ = 0,32). Сильнее всего увеличились обонятельная луковица и обонятельная кора. Это со всей очевидностью указывает на развитое обоняние. Полушария переднего мозга стали выпуклыми благодаря развитию неокортекса; они закрывают средний мозг и эпифиз при взгляде сверху, как у млекопитающих. Передний мозг Morganucodon шире мозжечка, хотя мозжечок тоже заметно вырос по сравнению с базальными цинодонтами.
Увеличение мозжечка говорит об улучшенной координации движений. На это же указывает и более толстый, чем у базальных цинодонтов, спинной мозг.
Развитие неокортекса у древних млекопитающих было связано в первую очередь с совершенствованием соматосенсорных функций (см. Somatosensory system). Значительная часть неокортекса у примитивных млекопитающих, таких как опоссум, — это так называемая соматосенсорная кора, которая отвечает за сбор и анализ сигналов, приходящих от многочисленных механорецепторов, рассеянных по всему телу. Особенно много таких рецепторов приурочено к волосяным фолликулам.
По мнению многих палеонтологов, волосы выполняли сначала тактильную (осязательную) функцию, а для терморегуляции стали использоваться позже, когда у предков млекопитающих начала развиваться гомойотермия (теплокровность). У Morganucodon и Hadrocodium достоверных остатков волосяного покрова пока не обнаружено, однако их близкий родственник — похожий на бобра маммалиаформ Castorocauda — был покрыт густым мехом, состоявшим, как у современных зверей, из осевых волос и подшерстка (см.: Скелет удивительного водоплавающего зверя обнаружен в юрских отложениях Китая, «Элементы», 12.03.2006). Это позволяет предположить, что Morganucodon и Hadrocodium тоже были покрыты шерстью. По мнению авторов, появление неокортекса у маммалиаформ было тесно связано с развитием волосяного покрова и осязания.

Мозг Morganucodon, базального представителя маммалиаформ, иллюстрирует первый этап прогрессивной эволюции мозга в ходе становления млекопитающих. На этом этапе увеличение мозга было обусловлено развитием обоняния, осязания и координации движений. Изменения в строении внутреннего уха свидетельствуют также о возможном улучшении слуха.
Hadrocodium, продвинутый представитель маммалиаформ и ближайший родственник «настоящих» млекопитающих, иллюстрирует второй этап развития мозга. Коэффициент энцефализации у Hadrocodium равен 0,5, то есть мозг увеличился еще в полтора раза по сравнению с Morganucodon и достиг размеров, характерных для некоторых настоящих млекопитающих. Мозг вырос в основном за счет обонятельных луковиц и обонятельной коры. Таким образом, второй этап прогрессивной эволюции мозга тоже был связан с развитием обоняния.
Косточки среднего уха (молоточек и наковальня) у Hadrocodium отделены от нижней челюсти, что является одним из главных определительных признаков млекопитающих. У Morganucodon, как и у всех остальных рептилий, эти косточки входят в состав нижней челюсти (см.: Новая палеонтологическая находка проливает свет на раннюю эволюцию млекопитающих, «Элементы», 17.03.2007). Авторы, однако, полагают, что отделение молоточка и наковальни от нижней челюсти вряд ли было связано с радикальным улучшением слуха, потому что строение внутреннего уха у Hadrocodium такое же, как у Morganucodon. Авторы также намекают, ссылаясь на данные по эмбриональному развитию опоссума, что это важное изменение черепа могло быть просто побочным следствием разрастания обонятельной коры переднего мозга.
Третий этап прогрессивной эволюции мозга соответствует переходу от высших маммалиаформ, таких как Hadrocodium, к настоящим млекопитающим. На этом этапе обоняние становится еще более тонким, о чём свидетельствуют специфические изменения решетчатой кости: на ней образуются носовые раковины, поддерживающие разросшийся обонятельный эпителий.
Новые данные показывают, что потребность в тонком чутье была, по-видимому, главным стимулом развития мозга в ходе становления млекопитающих. У млекопитающих, как известно, обоняние развито намного лучше, чем у всех остальных наземных позвоночных. Скорее всего, это изначально было связано с приспособлением к ночному образу жизни (см.: Обоняние и цветное зрение в эволюции млекопитающих развивались в противофазе, «Элементы», 18.06.2008). К концу триаса — началу юры синапсиды окончательно проиграли диапсидам конкуренцию за «дневные» ниши, и выжить удалось только тем из них, кто сумел «уйти в ночь», выработав совершенное обоняние для ориентации в потемках.
Источник: Timothy B. Rowe, Thomas E. Macrini, Zhe-Xi Luo. Fossil Evidence on Origin of the Mammalian Brain // Science. 2011. V. 332. P. 955–957.



